The requirements for general organic pigments used for polymer coloration, namely, wide acceptable temperature range, fine dispersion in the carrying material, lightness stability, a good migratory stability, are challenging, and result in high cost of organic pigments. At the same time, the supply of the raw materials for the synthetic pigments is limited by strict ecological and polymer coloration requirements. The microbial pigments being secondary metabolites are the diversified composition according to their structure and may be attributed to the ecologically safe compounds in the chem.- ical industry.
The development of products based on new biological chromophores is a novel aspect in biotechnology. Serratia marcescens synthesizes the membrane linked red pigment prodigiosin—pyrryldipyrrylmethene linked to the membrane [2,3]. Prodigiosin is a well known pigment and numerous factors have been reported to affect its production in the culture [4-7]. In this search, the economically simple culture medium on the base of the efficient, environmentally safe components for prodigiosin synthesis was used. The necessary bulk of biomass as the initial raw material for pigment isolation has been provided.
The present study aims at revealing the effect of the raw material quality on yield of pigment in the batch culture of Serratia marcescens and elaborating the method of the extraction from biomass met requirements for coloration of polyolefines by pigment preparation prodigiosin.
2. EXPERIMENTAL
2.1. Bacterial Strain and Culture Conditions
The pigmented strain Serratia marcescens strain 9986, in this study was obtained from the Culture Collection of Kazan Federal University (Russia). The culture was maintained in the liquid medium [5] and also in broth containing meat peptone (5 g·l−1) and glycerol (10 g·l−1) [2] as a standard medium for cultivation. The strain was cultivated in flasks 250 ml (total of 50 ml), pH 7. 2, at 28˚C, at 180 rpm on a shaker (IS-971 R IEIO Tech., Korea) in the darkness. The culture was grown in flasks 2.0 l containing 1, 1 l growth medium at the same conditions of the incubation for accumulation of pigmented biomass under controlled factors of the fermentation. It was also grown in two variants on the medium with meat peptone (5 g·l−1) (Sigma) or with peptone enzymatic digest from soybean (5 g·l−1) (Fluka).
Cell growth was determined at 670 nm (OD) so that the optical density of growing culture and spectrum of prodigiosin (535 nm) have been divided.
The experiments were repeated three times and measurements were made for each time point of the incubation.
2.2. Prodigiosin Essay
To extract prodigiosin 1.0 ml of cultural liquid from grown cells or from cell suspension to 9.0 ml of acidified ethanol (10 ml of ethanol and 1 ml of 1 N HCl) added. Time of extraction made 20-min, the cell debris was removed by centrifugation at 9200 g, for 10 min, 5˚C. The prodigiosin concentration was measured spectrophotometrically at 535 nm in the growing culture and in the cellular suspension on the scanning spectrophotometer a Lambda-25 (Perkin Elmer, USA). Pigment concentration was calculated using extinction coefficient for prodigiosin (51.5 × 103 l·g·cm) [4].
Definition the Size of the Pigment Protein Particles
The size of the pigment protein particles was determined to the method of dynamic light scattering (DLS) [8].
2.3. Treatment of Biomass for Prodigiosin Preparation
The method of the extraction from biomass of pigment preparation prodigiosin was elaborated according to technology of dyeing polyolefines. The grown pigmented culture was subjected to centrifugation at 6000 g, for 15 min, 5˚C. In the crude pigmented biomass the detergents sodium dodecylsulphate (SDS) (0.1 - 0.5 wt%) or nonionic Triton X-100 (10.0 mM) were introduced. The solubilizable cell suspension dissolved in 50 mM phosphate buffer (pH 7.0) was tested on the lightfastness of prodigiosin by exposing the pigment suspension under lighting the fluorescent lamp (Hitachi) at a flux of 45.0 µM photons m−2·s−1 until 48 h (Table 1). In the elaborated method the treatment of the crude biomass by 0.1 wt% SDS was undertaken. The solubilization of the pigmented biomass by SDS is a necessary stage for later use. Then an effective extraction of the pigment preparation from solubilizable biomass by polar organic solvent ethanol no less than 4 - 5 times was consisted Pigment alcohol solution was concentrated in the rotary vacuum evaporator under temperature 62.0˚C - 63.0˚C.
The production of the coloration polymeric compositions by pigment preparation prodogiosin was described [9,10].

Table 1. Lightfastness of the pigment suspension treated by detergents (%).
2.4. Dyeing of Polyolefines by the Prodigiosin Preparation
The bright red prodigiosin colour of the concentrates and the masterbatches were achieved by the method of polymer dyeing in the Central Laboratiry of J-SCo “Kazanorgsyntese” [11]. In this study the polyethylene and the ultrathene in granules were applied for coloring elaboration. The manufacture of the polymer compositions was carried out on the press (Polymix 110 P, Schwabenthan, Germany) by introduction granular polymer. The process of rolling was at: the temperature 150˚C - 155˚C on the front roller and 135˚C - 140˚C on the back roller. The time of rolling process was 20 - 25 min. The pigment suspension was introduced gradually up to rolled polymer sheet for the equilibrium coloration of the dyed-stuff. The manufactured polymer sheet served as a colored concentrate (the colour masterbatches additives). The press for compressing various thickness of dyed samples was used (Fantune, Holland). The cutting out was applied for the preparation of dyed samples in the process tests of their quality (CELST, Italy). Dyed form of molded plates-dimensions of 20.0 × 20.0 mm and the thickness of 1.0 ± 0.1 mm were also evaluated according to the technical standard.
The pigment preparation prodigiosin was used in the technology dyeing as a thick suspension-concentrate without spraying in the air.
3. RESULTS AND DISCUSSION
3.1. Cell Growth and Prodigiosin Production Conditions
In this elaboration the success of a biotechnological process for exploitation at industrial level of prodigiosin has been revealed. It was found the novelty of the realizable advantages associated with more economical consumption of prodigiosin preparation in the dyeing polyethylene concentrates, a happy choice the conditions for extraction pigment from biomass useful to the colorant technology, the brief time of fermentation etc.
The biosynthesis of prodigiosin by pigmented strain of Serratia marcescens under aerobic conditions in the batch culture is realized. The pigment production is highly variable among species Serratia and it depends on many factors such as the specific strain, the temperature, the incubation time, the illumination, the pH medium and others [2,5,12]. The illumination conditions (light/darkness) affected both the biosynthesis of prodigiosin and energetic metabolism of the pigmented strain ATCC 9986 Serratia marcescens were shown [8].
The phenomenon of prodigiosin production was discovered in the batch culture at temperature 28˚C. Moreover, the content of prodigiosin in the culture grown in darkness was higher than that grown in light. The pigment biosynthesis beginning from 18 - 20 hours of incubation of culture was revealed. A maximum of pigment synthesis in the culture was reached at the end of exponential growth phase and initial stationary phase within 50 - 54 hours of the incubation that corresponded to the stage of active cell growth (Figure 1). In this way the time of fermentation is mostly preferably for pigment isolation from fermented broth. However, the accumulation of prodigiosin in the culture growing in darkness was higher than that grown in the light, 42.0 - 60.0 µg·ml−1 and 30.0 - 36.0 µg·ml−1, respectively. Dark culture retained its bright red color to a dark cherry one up to 7 - 9 days and the pigmented cells had been already lysed.
Serratia marcescens, like other Enterobacteriaceae, grow well on ordinary media and other differential and selective media [13-15]. The regular liquid medium currently being used for prodigiosin biosynthesis is nutrient meat peptone-glycerol broth. It was used as a growth medium for Serratia marcescens strain 9986 where the glycerol was the carbon source and a peptone was as the nitrogen source. The cheap medium peptone digest from soybean as economically cheap source was used also in the nutrition medium. Figure 2 shows the culture growth
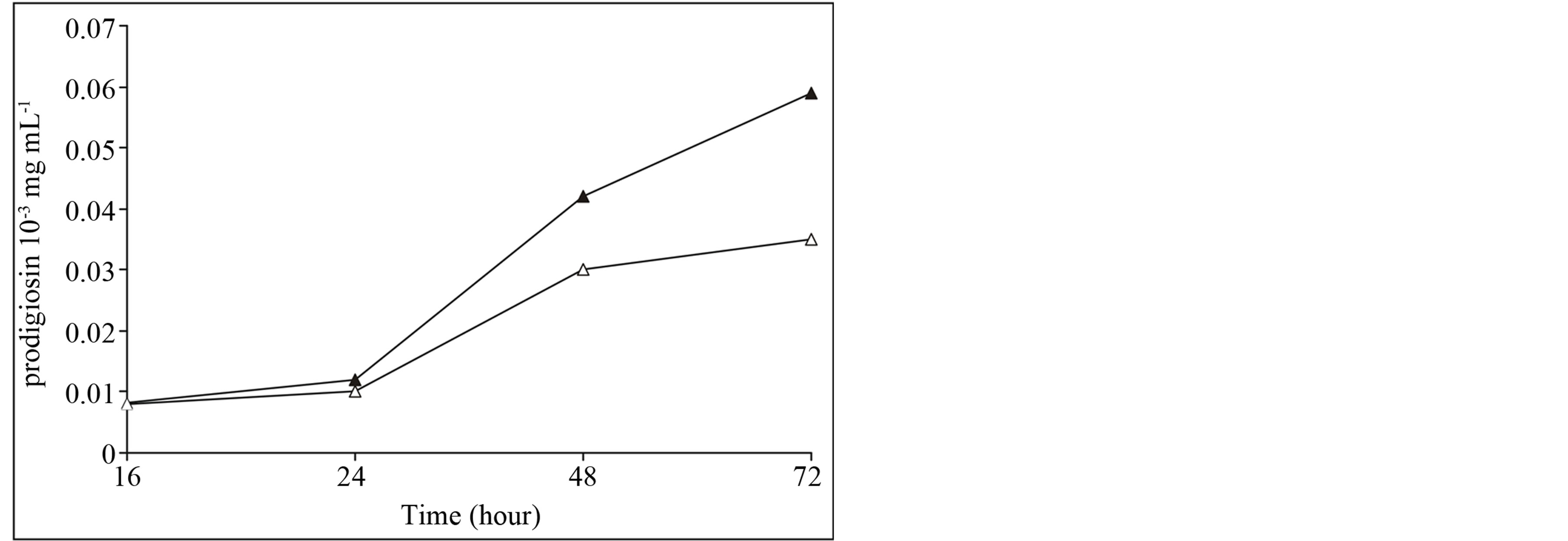
Figure 1. The biosynthesis of prodigiosin in the light (¡), in darkness (l) Serratia marcescens strain 9986 was incubated in broth containing meat peptone and glycerol. The pigment synthesis (E 535 nm) in darkness was higher by comparison in the light 42.0 - 60.0 µg·ml−1.
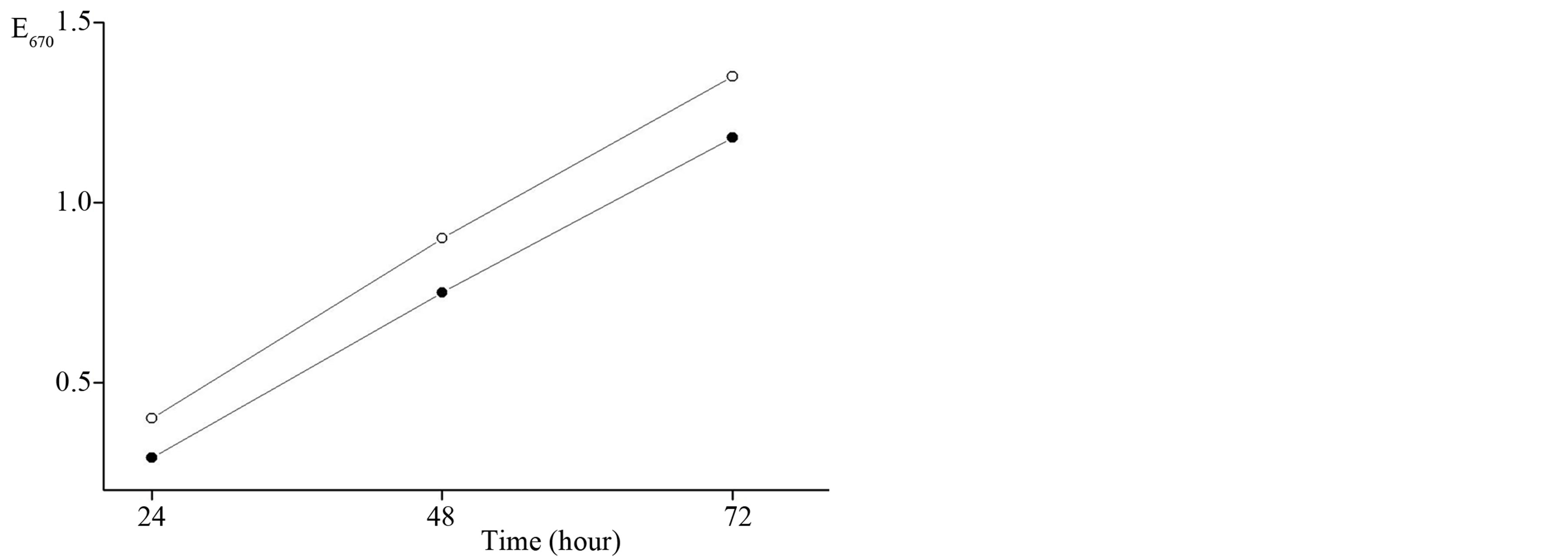
Figure 2. The growth of Serratia marcescens in the broth meat peptone (¡), on soybean peptone (l) a biomass in the active growth phase of Serratia marcescens strain 9986 on both media was not differed statistically significant.
on a peptone meat and peptone digest soybean up to 72 h of the incubation. The accumulation of a biomass in the active growth phase by 48 h on both media differed insignificantly. In the initial stationary phase the established distinctions in biomass were not statistically significant. The analogous correlation was observed in the pigment biosynthesis since a low price of peptone soybean as a vegetable raw material was suitable fully for fermentation and pigment biosynthesis.
There is the correlation of the pigment accumulation and pH of the culture medium. As shown in Figure 3 in the first 24 h of incubation the culture growth on both media was accompanied by a decrease of pH 6.5 - 6.6. Subsequently, the alkalinity of the media during the all course of growth was up to pH 7.5 - 7.0 and pH 7.7 -7.2 on peptones meat and from soybean, respectively. Therefore, pH nutrient medium is one of the determining factors in the technology of pigment biosynthesis [16].
Determination the Size of the Pigment Protein Particles
The hydrodynamic diameter of pigment protein complex was revealed by the method of the dynamic light scattering (DLS). The hydrodynamic diameter of the pigment protein samples within the limits of 7.5 - 8.0 nm was shown (Figure 4). In dependence on protein structure and density the globular proteins of such size of the molecules could be characterized by molecular mass 100 - 200 KDa [17]. Our results could be interpreted as a complex of pigment and protein by molecular mass a little smaller than 100 KDa basing on results of DLS. The revealed complex consisting from particles of diameter 7.5 - 8.0 nm was in a lower range limits. The results of DLS also showed that the surfactant sample was characterized by the mean hydrodynamic diameter (DH) of particles about 6 nm which come to an agreement with
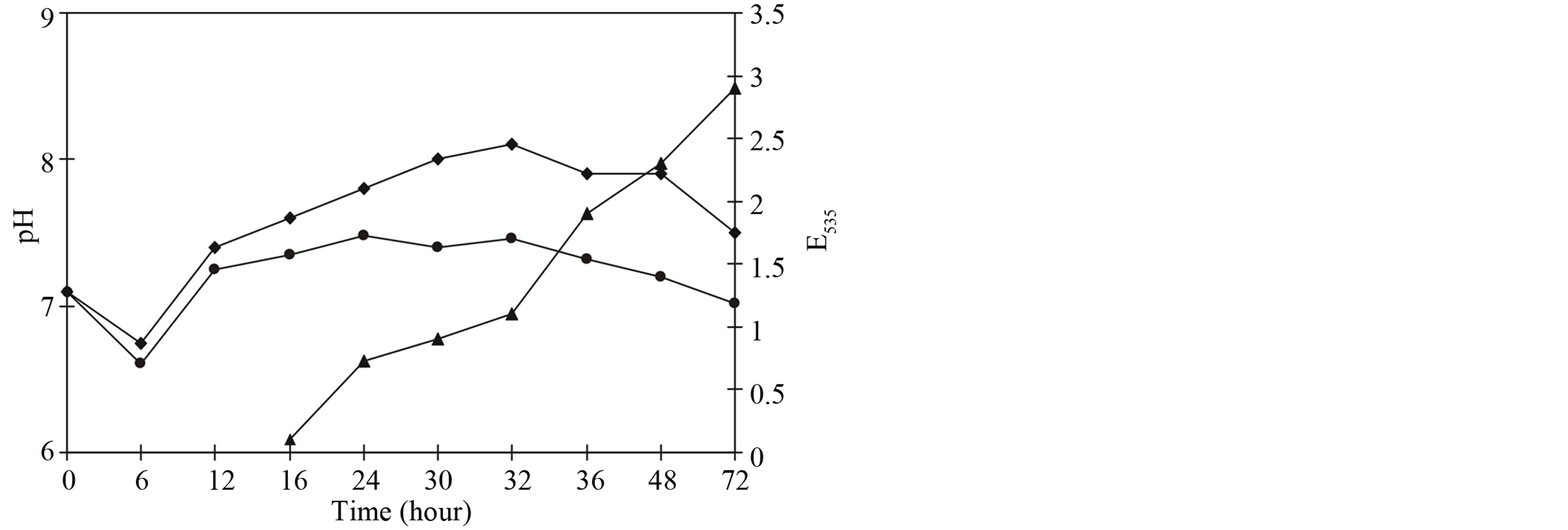
Figure 3. The biosynthesis of prodigiosin and the course of pH in the broth meat peptone (l), soybean peptone (♦), prodigiosin (▲) In the first hours of the incubation of Serratia marcescens pH of the media were descended up to 6.5 - 6.6. Thereupon pH in the growing culture is up to 7.5 - 7.0 and 7.7 - 7.2 on peptone meat and soybean, respectively. Optimal pigment biosynthesis (▲) depended on pH nutrient medium.
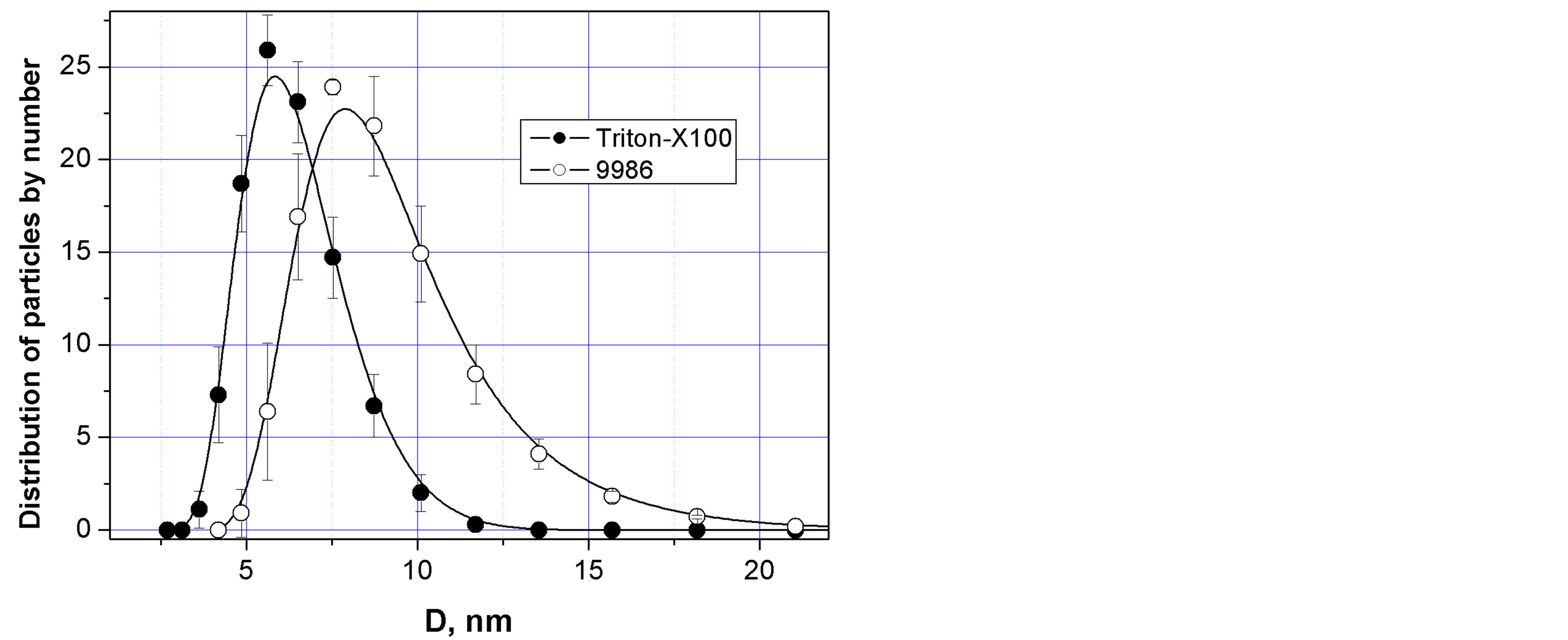
Figure 4. Mean hydrodynamic diameter (nm) of particles isolated from Serratia marcescens strain 9986. Triton X-100 (l) and pigment protein complex (¡). Pigmented cells were suspended in 10 mM Triton-X100 and the lysate was centrifuged at 9200 g. The supernatant was saturated by (NH4)2SO4—0,2 N (saturation). The colored of pigment protein fraction was resuspended in the bidisllate. The hydrodynamic size of pigment protein particles (¡) was measured (by method of dynamic light scatterning (DLS), Malvern Instruments Ltd., Malvern, UK) and the size of micelles of Triton X100 (l).
the size of micelles of Triton X-100 as was shown in literature [18,19].
3.2. Preparation of Prodigiosin from Biomass
As it was seen earlier, Serratia marcescens determines pigmentation in correlation with growth conditions namely carbon source and nitrogen, the regular variation of pH in the whole period of fermentation, temperature, illumination etc. For strain 9986 of Serratia marcescens the optimal biosynthesis of prodigiosin was observed by 50 - 54 hours of the incubation and at take one’s temperature 28˚C. The pigment yield 0.2 - 0.4 mg·l−1 from culture medium was obtained on a meat peptone-glycerol broth in the batch culture under aerobic conditions. Next essential stage of the downstream procedure was focused on the elaboration of the pigment extraction from biomass to meet technological requirements of the polymer coloration.
The technology of pigment isolation consisted from the treatment of crude biomass by detergents. Originally the growing culture at 50 - 54 hours of incubation was sedimented by centrifugation to remove the cell debris. The detergent ionic SDS and the nonionic Triton X-100 for the crude biomass to screen the extractive biomass were added [15,20,21]. After regular distribution during half an hour, the solubilizable pigment suspension was tested on the lightfastness of prodigiosin. The selected ionic surfactant SDS was more effective to the lightfastness of pigment by comparison with Triton X-100 (Table 1). Thus, ionic detergent SDS in the concentration of 0.1% - 0.5% to the crude biomass was used. Less concentration of surfactant SDS (<0.1%) was caused by the loss of the decoloration prodigiosin in the polyethylene concentrate. The increased content of SDS more 0.5% in the technological treatment of polymer concentrate resulted in a considerable worsening of the colored polyethylene. The polymer sheet provoked the slide in the rolling process. The offered elaboration is favoured using 0, 1% of SDS in the technological process gains thermostability of the colored polyethylene sheet and the concentrate.
3.3. Working of Dyeing the Polyolefines by Prodigiosin
Prodigiosin pigment preparation as a thin suspension (paste) was introduced available to polyethylene concentrate. This was conducived to negligible quantity aglomerates in the polymer ware. The molecular mass of the introduced pigment in the polyethylene was smaller than 100 KDa as shown [8].
According to the technological method the colored concentrate 4.0% - 8.0% mass was introduced into the melted down polymer. The time of rolling process was 20 - 25 min. The manufactured polyethylene concentrates and the colored polyethylene concentrate by prodigiosin preparation were conformed to the technological tests (Table 2). Dielectric tests of colored masterbatches corresponded to the experimental data and technical standards (Table 3). Besides in the process of manufacture, the pigmented spicemens of the polymer concentrates have been painted uniformely by prodigiosin and
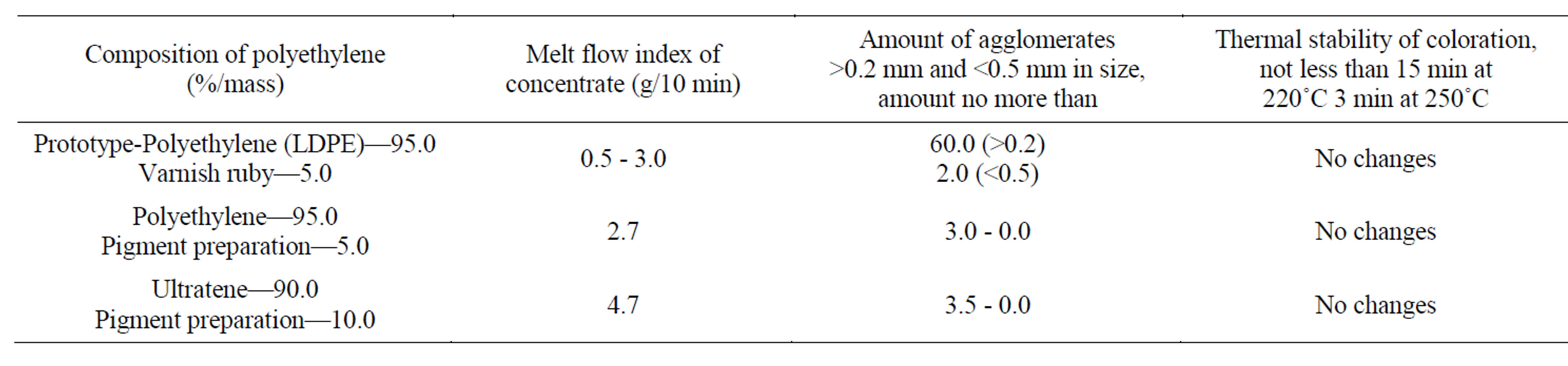
Table 2. Tests with colored polyethylene masterbatches.
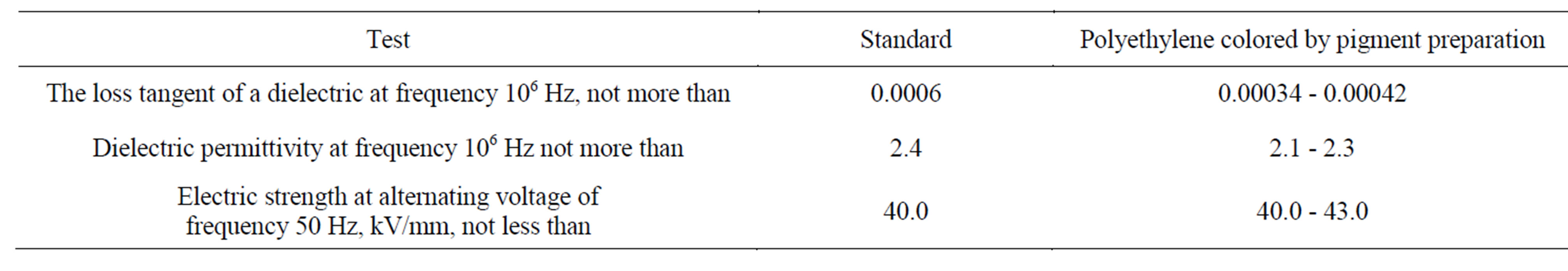
Table 3. Dielectric tests with colored polyethylene masterbatches.
the migratory capacity of pigment from polymer was absent during some years. According to the physical, mechanical parameters and dielectric properties the masterbatches met the standard requirements of cable materials. These results indicate that according to elaborated method the prodigiosin preparation can be considered as a novel type of biological dye-stuff.
The results from Table 4 showed that prodigiosin content in the coloring polymer was very low (mg % mass) than in application to synthetic organic dyes utilizing in the chemical industry. This advantage in the colored polymer results in its minimal influence on the technological properties of the dyeing concentrate and masterbatches (thermal stability, dielectric etc.).
This search is related to prodigiosin red pigment and based on the current state of our knowledge. In the present industry the red polyethylene masterbatches include high toxic dyes. New resources in the technology for coloration of polymers with the ecologically safe microbe pigments have offered the following radically novel aspects. It is essentially important to note that prodigiosin in the process of fermentation is synthetised as microbe metabolite unlike the organic dyes the synthesis of which is the multiple stage process. The pigments of microorganisms in their composition belong to derivatives of the ordinary metabolites or cellular components and they are ecologically safety compounds which offer principal advantage over other synthetic dye-stuff. However, the microbe pigments being secondary metabolites are synthesized and produced maximum in the period of the stationary growth phase and further they are not metabolized. Serratia marcescens strain 9986 is a producer of prodigiosin bright red pigment which was used as a dye of polyethylene. The fermentation process of microbial strain and the pigment biosynthesis are car-
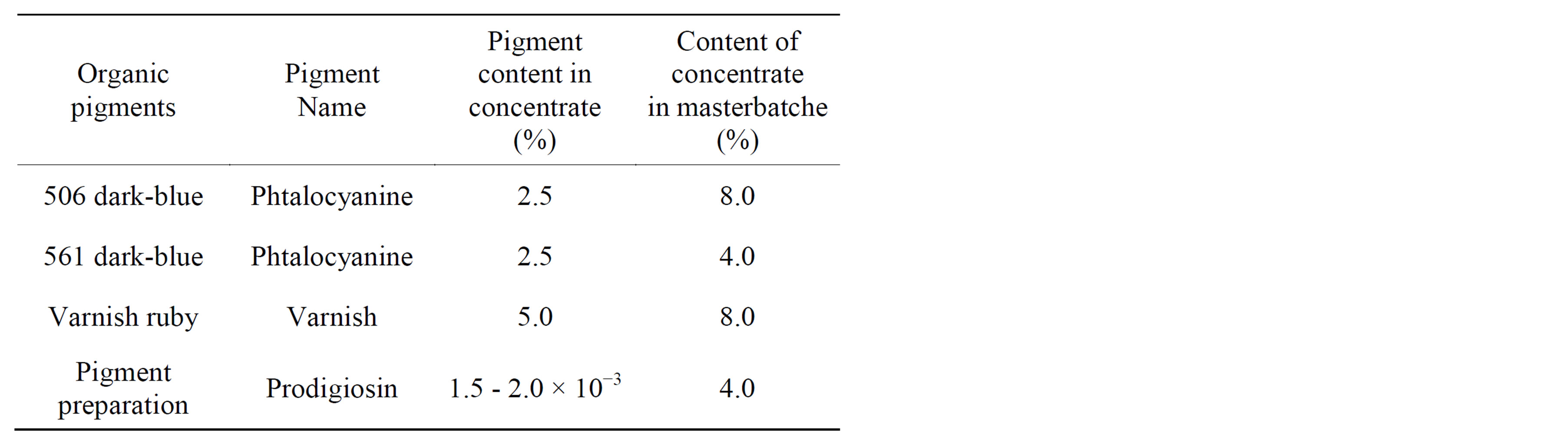
Table 4. Consumption of pigment concentrates in the colored polyethylene.
ried out under the strictly controlled conditions. The period of fermentation is shorter in comparison with other biological producers (fungi, algae). The consumption of pigment preparation is more economical in the technology of the dyeing of polymer concentrates. The lower pigment content in the colored concentrates and masterbatche renders smaller influence upon technological parameters of polymer. Other advantages of these method excepting of the rapid cell growth, inexpensive culture media, and a fermentation process that can be easily scaled up to produce industrial quantities of the microbe pigment from target product. In addition, the elaborated method for coloration of polyolefines described here is more effective with minimal effect on an environment than the existing methods in application to organic dyes.
4. CONCLUSION
The biosynthesis of prodigiosin was closely connected with growth parameters which were necessary and intended correctly. A pigment yield 0.2 - 0.4 mg·l−1 from culture medium was obtained on a meat peptone-glycerol broth in the batch culture under the aerobic conditions. The elaborated method of the coloration of polyethylene by prodigiosin preparation allowed carrying out the articles of common use complying with the technological standard. The assortment and the application of novel polyethylene concentrates and masterbatches based on the coloration by bacterial pigment are expanded including raw material supply and appropriate innovations in the chemical industry.
ACKNOWLEDGEMENTS
This work was support. by firm J-S Co Kazanorgsynthes. The authors would like to thank chief technologist N. N. Valiyeva for her helpful technological elaboration and insightful discussion of obtained technological results.
NOTES