Received 1 June 2016; accepted 19 July 2016; published 22 July 2016

1. Introduction
Agarwood is a resinous substance occurring in Aquilaria trees of Thymelaeceae family that is found in the foothills of Himalayas to the rain forests of Papua New Guinea. The tree has the unique feature of producing resin- impregnated heartwood which is fragrant and economically highly valued depending on the oleoresin content of the wood. The wood is used to manufacture some of the medicines, incense and perfumes across Asia and Europe. The agarwood is reputed to be the most expensive wood in the world and in the consumer countries it ranged from a few dollars per kg for low quality material to more than US$30,000 per kg of top quality wood [1] . The formation of agarwood is mainly attributed to the defense reaction of trees either physically or chemically when they are exposed to biotic and abiotic stresses [2] - [4] . The agarwood causal agents so far studied as physical [5] , chemical and biological agents [6] . The form of mechanical injury is considered as physical inducer of agarwood formation while the induction of chemicals, viz. oil, sugar and methyl jasmonate are reserved as chemical inducers. However, the quality and quantity of agarwood formation were not found to be greater in case of the physical and chemical inducers. Later, the studies on the biological agents in rainforest areas on the infection site of the agarwood trees revealed that the fungal microbes could be the potential agents to induce the formation of agarwood [7] [8] . As a response to the fungal infection, the tree produces a high resin in volatile organic compounds that aids in suppressing or retarding the growth of the fungus. As the fungi caused injury to the tree trunk, the tree underwent several biochemical reactions and produced a white, milky substance called oleoresin. Once the production of aromatic trunk or agarwood is complete, the tree slowly starts drying up, signaling its readiness to be harvested. From this, it is understood that resin wood or agarwood is the result of oleoresin accumulation in response to fungal infection. However, little information is available on the potential fungi and its enzymatic activities that are associated with the formation of agarwood. In this circumstance, it is believed that the study on isolation and characterization of fungi associated with agarwood formation could be the first step in proceeding further research work on the standardization and development of artificial inoculation agents so as to produce the high quality agarwood. With this background information, the current study was carried out with the objectives: 1) identification of fungi from naturally infected agarwood; 2) study of the enzymatic activities of agar wood fungi.
2. Materials and Methods
2.1. Collection and Isolation of Agarwood Fungi
The infected agarwood samples were collected from Dibrugarh, Hojai and Rain Forest Research Institute, Jorhat of Assam State and Sarkkapathi, Aliyar Nagar of Tamil Nadu State and Kumily, Idukki District of Kerala State in India. The wood samples were chipped into 1 cm2 and used for the isolation of fungal microorganisms. The cut chips were surface sterilized in 0.1% mercury chloride for 30 Sec and rinsed in sterile distilled water for three times and placed on PDA medium prepared in Petri dishes and incubated at room temperature (26˚C ± 2˚C) for 7 - 14 days. The fungal colonies appeared on PDA plates were sub-cultured and an axenic culture of the fungi was obtained by single hyphal tip technique and maintained in PDA slants to carry out further characterization studies.
2.2. Study on Morphological Characters of Fungi
The pure culture of individual fungi was observed for mycelial colour, texture and growth characters [9] . The microscopic slides were prepared from individual cultures to observe spores and mycelia under microscope. Based on the colony characters and morphology, the fungal isolates were tentatively identified.
2.3. Assay of Cellulase Activity of Agarwood Fungi
The estimation of cellulose degradation was carried out as described in Gajera et al., 2008 [10] . The fungal cultures were grown in petri plates containing PDA. After eight days of inoculation, 10ml of sterile distilled water was added to each of the petri dishes and the mycelium was sufficiently wet. The mycelium from the petri dishes were transferred to conical flask and shaken well in orbital shaker at 120 RPM at 28˚C for about 4 hours. After that it was transferred to 30 ml centrifuge tubes and centrifuged at 10,000 RPM for 10 min. The culture (0.2 ml) was incubated with 0.4 ml of 0.1 M sodium citrate buffer (pH 5.0) containing 1% Carboxy Methyl Cellulose (CMC). After 30 min of incubation at 55˚C, the glucose released was estimated by DNS method. The activity of cellulase was expressed in μmols∙min−1∙litre−1. The experiment was repeated with three replications for each isolate and the mean values were calculated.
2.4. Assay of Ligninolytic Enzyme Activity of Agarwood Fungi
The activity of lignin modifying enzymes was detected by dye staining of lignin modifying basal medium (LBM) (KH2PO4: 1.00 g; Yeast Extract: 0.01 g; C4H12N2O6: 0.5 g; CuSO4∙5H2O: 0.001 g; MgSO4∙7H2O: 0.5 g; Fe(SO4)3: 0.001 g; CaCl2∙2H2O: 0.01 g per litre). LBM was supplemented with 1.6% w/v agar and autoclaved. To 100 ml of LBM, 1 ml of 20% aqueous glucose solution and 1 ml of 1% w/v aqueous tannic acid solution was added. After sterilization, the medium was transferred aseptically into the petri plates and used for the assessment of lignin degrading enzyme activity of the fungal isolates. Eight days old fungal culture was inoculated on LBM medium and the plates were incubated at 25˚C in darkness. The plates were examined regularly up to ten days. The lignin degrading enzyme activity was recorded based on the appearance of brown oxidation zones around the colony [11] .
2.5. Assay of Laccase Activity
The fungal cultures were grown in Petri plates containing PDA. After eight days of inoculation, 10ml of sterile distilled water was added to each of the petri dishes and the mycelium was sufficiently wet. The mycelium from the petri dishes were transferred to conical flask and shaken well in orbital shaker at 120 RPM at 28˚C for about 4 hours. After that, it was transferred to 30 ml centrifuge tubes and centrifuged at 10,000 RPM for 10 min. The supernatant was used for the assay of laccase. There action mixture consisted of 3 ml acetate buffer, 1 ml guaiacol and 1 ml enzyme source. The blank sample was prepared using 1 ml of deionized distilled water instead of enzyme source. The mixture was incubated at 28˚C for 5 min and absorbance was read at 450 nm in UV spectrophotometer. Enzyme activity was expressed as International Units (IU), where 1 IU is defined as amount of enzyme required to oxidize 1 micromole of guaiacol per min. The laccase activity in Uml−1 is calculated from the extinction coefficient of guaiacol (6.39 M−1∙cm−1) at 450 nm by the formula:
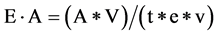
where E∙A = Enzyme Activity (Uml−1), A = Absorbance at 450 nm, V = Total volume of reaction mixture (ml), v = enzyme volume (ml), t = Incubation time (min) and e = Extinction Coefficient (M−1∙cm−1) [12] . The experiment was repeated with three replications for each fungus and the mean values were presented.
3. Identification of Agarwood Fungi Using PCR
3.1. DNA Extraction from Agarwood Fungi
DNA was extracted from the mycelial mat of each strain grown on PDB for 7 days at 28˚C by CTAB method described by O’Donnel et al. [13] and Srinivasa et al. [14] . The mycelium was thoroughly ground into a fine homogenate using pallet pestle mixer. Seven hundred μl of CTAB buffer [2% CTAB (hexadecyltrimethylammonium bromide) 100 mM Tris-HCl [pH = 8] 20 mM EDTA, 1.4 M NaCl 0.2% β-mercaptoethanol [added just before use] 0.1 mg∙ml−1 proteinase K [added just before use] was added to the tubes, mixed well and vortexed. This mixer was incubated at 65˚C for 10 minutes. Afterwards 400 μl extraction buffer (chloroform: isoamyl alcohol, 24:1, v/v/v) was added followed by gentle stirring on a vortex mixer till emulsion formation. The emulsion was centrifuged at 10,000 RPM for 10 minutes in a microcentrifuge at room temperature. An aliquot of the upper aqueous layer (600 μl) was collected, mixed with an equal volume of extraction buffer and re-centrifuged at 10,000 RPM for 10 min. From that upper layer 300 μl was taken and 0.5 volume of (150 μl) of 5 M NaCl and 600 μl of ice cold is opropanol was added and incubated at −20˚C for overnight. The mixture was centrifuged at 14,000 RPM for 10 minutes at 4˚C and the pellet was collected and suspended in 70% ethanol and re-centri- fuged at 5000 RPM for 10 min. Finally, the pellet was air-dried for 10 - 15 minutes and resuspended in 100 μl of Tris-EDTA (TE) buffer having 10 mM Tris-HCl (pH 8.0) 1 mM EDTA. To this, 2 μl RNAase was added and incubated at 37˚C for 90 min. The DNA concentration was estimated in DNA photometer and further confirmed by gel electrophoresis in 0.8% agarose gel containing 0.05% ethidium bromide.
3.2. PCR Amplification Using ITS Primers
The Internal Transcribed Spacers (ITS) primers ITS1 5’(TCCGTAGGTGAACCTGCGG)3’ as forward primer and ITS2 5’(GCTGCGTTCTTCATCGATGC)3’ as reverse primer were used for PCR amplification studies [15] . The PCR reaction was carried out in 50 μl volume. The PCR reaction contained 1 μl template DNA (100 ng∙μl−1) from agarwood fungi, 1 μl (4.5 pMol) of respective primers, 5 μl Red eye master mix which contains 1 μl dNTPs (10 mM dNTP stock), 5 μl PCR buffer (TrisHCl buffer), 5 μl MgCl2 (25 mM stock), 2.5 μl gelatin (1%) and taqpolymerase 0.5 μl of (5 units∙μl−1). Deionized water was used to make the total reaction volume up to 50 μl. In order to ensure the reproducibility of the reaction, appropriate negative controls (without DNA template) were also run.PCR tubes were placed in thermocycler (Eppendorf) for DNA amplification with following PCR program: Initial denaturation at 94˚C for 4 minutes, forty cycles of 94˚C for 1 minute, 36˚C for 1 minute and 72˚C for 2 minutes. Following the cycling, mixture was incubated at 72˚C for 10 minutes and then kept at 4˚C for 12 hours. The PCR products were resolved on 2% agarose at 50 V stained with ethidium bromide (0.5 μg∙ml−1) and photographed using gel documentation system (Alpha Innotech Corporation, USA).
Of several isolates amplified, the PCR product from the fungal isolate AR13 was purified from reaction mixture by agarose (1.2%, w/v) gel electrophoresis in TBE buffer containing 0.5 μg of ethidium bromide per ml. A small agarose slice containing the band of interest (observed under long-wavelength [312-nm] UV light) was excised from the gel and purified by using a QIA quick gel extraction kit (Qiagen, Inc., Chatsworth, California) according to the supplier’s instructions. This purification was performed to remove primer dimers and other residues from the PCR amplification. PCR product was sequenced at Chromous Biotech Pvt. Ltd., Bangalore, India. PCR sequence was identified using the basic local alignment search tool and submitted in GenBank nucleotide data bank, National Center for Biotechnology Information, Bethesda, MD, USA (http://www.ncbi.nlm.nih.gov/) and accession number was obtained for the isolate AR13.
4. Results
4.1. Morphological Identification of Agarwood Fungi
A total of 17 fungal cultures were isolated from agarwood infected samples collected from Dibrugarh, Hojai and Rain Forest Research Institute, Jorhat of Assam, Sarkkapathi, Aliyar Nagar, Tamil Nadu and Kumily, Idukki District of Kerala, India. Of these, 3 isolates were from Dibrugarh (DIB1, DIB2, DIB3) and 4 isolates from Jorhat (RFRI4, RFRI5, RFRI6, RFRI7) of Assam, 6 isolates from Tamil Nadu (AR10, AR11, AR12, AR13, AR14, AR15) and 4 isolates from Kerala (K1, K2, K3 and K4). The colony characters such as colour of the mycelia mat, texture, mycelia growth of individual fungal isolates were observed on PDA medium. The mycelium and conidial characters of the individual fungal isolates were examined under microscope. The observations on colony characters and conidial characters were aided in tentative identification of the fungal cultures (Table 1). The fungi were identified as Aspergillus, Penicillium, Fusarium, Lasiodiploidia and Chaetomium.
4.2. Enzymatic Activities of Agarwood Fungi
A significantly higher activity of cellulase was measured in Aspergillus isolate AR13 (1181.952 μmole∙L−1∙min) isolated from Aliyar Nagar of Tamil Nadu followed by Aspergillus isolate K1 (1140.608 μmole∙L−1∙min) whereas the lesser activity was observed in Chaetomium isolate K4 (449.92 μmole∙L−1∙min) (Table 2).
Laccase activity of fungal isolates revealed the higher secretion of laccase enzyme by Aspergillus isolates K1 (18.116 Uml−1) and AR13 (17.125 Uml−1). These two isolates showed significantly greater activity of laccase production when compared to the other isolates of agarwood fungi (Table 2).
The results of lignin degradation in LME medium supplemented with tannic acid showed brown layer as indicator of positive reaction for lignin degradation. Of seventeen isolates, Aspergillus isolates AR13 and K1 showed stronger ligninolytic activity. The isolates of Fusarium DIB2, RFRI7 showed the medium activity whereas less activity was observed in Lasiodiploidia DIB3, Chaetomium AR11 and Aspergillus K1 (Table 2).
4.3. PCR Amplification of ITS Region of Agarwood Fungi
Out of 17 isolates used in the PCR studies, the isolates RFRI4 (Lasiodiploidia sp.), AR10 (Chaetomium sp.), AR11 (Chaetomium sp.), AR14 (Penicillium sp.) and AR13 and K3 (Aspergillus sp.) have amplified ITS1 and ITS2 primers. The DNA gel electrophoresis showed that the amplified products were around 560 bp. Of this, the PCR product amplified from the isolate AR13 (Aspergillus sp.) was purified and sequenced (Figure 1). The nucleotide sequence data showed 93% homology to Aspergillus genus when blast with NCBI database. The nucleotide sequence of agarwood fungal isolate AR13 was submitted to NCBI and assigned with Accession No. KF263963.
![]()
Table 1. Morphological characters of agarwood fungi.
![]()
Figure 1. PCR amplification of ITS region of Aspergillus isolate AR13. Lane M: 100 bp marker. AR13: Isolate AR13.
![]()
Table 2. Enzymatic activities of agarwood fungi.
1Note: slope value = 7296. 2Note: Extinction coefficient for Guiacol 6.39. **Enzyme activity is significantly different from the other fungal microbes.
5. Discussion
The current study identified the colonization of agarwood by Aspergillus, Penicillium, Fusarium, Lasiodiploidia and Chaetomium fungi. Previous studies have also showed that fungal species such as Aspergillus sp., Botryodiplodia sp. (Lasiodiplodia sp.), Diplodia sp., Fusarium bulbiferum, F. laterium, F. oxysporum, Penicillium sp., Pythium sp. and Trichoderma sp., could infect Aquilaria spp. [8] [16] . The major enzyme activities were observed in Aspergillus isolate AR13 in the present study. This could favour the potential infection of agarwood by Aspergillus fungus. It was also reported in Wiriadinata [17] that agarwood had mainly been attacked by opportunistic fungi from soils of rainforest environment. It is regarded as a weak pathogen and does not cause heavy damage to trees. However, the continuous release of enzymes by this fungus could favour defense mechanism in agarwood. Thus, the enzymes involved in the pathogenesis process were assayed in the current study, of which, Cellulose is the major framework molecule of the plant cell wall existing as microfibrils with matrix molecules (glycoproteins, hemicelluloses, pectins, lignins) filling the spaces between the microfibrils and cellulose chains. Cellulolytic enzymes produced by fungi play a role in softening and disintegrating plant cell walls [11] . Thus all 17 fungal isolates were assessed for cellulase activity by dyediffusion method. The study revealed the higher cellulolytic activity by Aspergillus isolate AR13 when compared to all other isolates. Similarly, lignin is a highly branched heterogeneous polymer found principally in the secondary cell walls of plants, although primary walls can also become lignified. In the current study, all seventeen fungi were showed the lignin degrading activity. The presence of lignin degradation was verified based on the brown oxidation zone present around the colony where in the isolate AR13 (Aspergillus niger) showed higher activity. Similarly, the laccase activity was recorded higher in Aspergillus niger isolate AR 13 in the current study. The pathogenesis process by fungus is determined by the production of laccase activity [18] . Similar results were obtained by Desai et al. [12] where Trichoderma genus produced higher levels of laccase activity under solid state condition.
From the current study, it is believed that the formation of agarwood could be a result of plant defense mechanism towards fungal attacks by producing resinous compounds as secondary metabolite [3] [19] [20] . It is also assumed in the current study that the series of enzymes produced by the pathogenic or saprophytic fungal microorganisms could play a major role in defense mechanism which ultimately becomes responsible for agarwood production. In this context, it is possible to develop Aspergillus niger AR13 as artificial agent to inoculate the agarwood.
Interestingly, two of our isolates from our collection i.e. Aspergillus and Lasiodiplodia were also reported as endophytic fungi in Mitra and Gogoi [21] . In this context, our cultures should be tested for their endophytic nature and their role in agarwood formation. Although the possibility of inducing agarwood formation by deliberately infecting trees with fungal isolates has been postulated to induce resin, it has to be tested at various agroforestry settings having different environmental factors and weather conditions. Further, the isolate AR13 which is to be tested for agar formation studies should be verified for its host specificity. It is otherwise should be proved as non-pathogenic to other crop plants.
NOTES
![]()
*Corresponding author.