The uptake of primary (N, P, K) and secondary (Ca, Mg) macronutrients in different plant organs greatly depend on fauna activities presents in the rhizosphere soils together with the capacity of the soil to refill in the given soil solutions [5]. Primarily, N is necessary for the formation of amino acids which are building blocks of protein also aid for plant growth which triggered through cell division. Besides N is directly involved in photosynthesis and is a necessary component of several vitamins in legumes [1]. The demand for N in a deficient soil is normally achieved by the use of chemical fertilizers. However, the high cost of mineral N fertilizers may be counteracted by other alternative means to meet the N demand through the use of beneficial bacteria such as Rhizobium [6].
Phosphorus (P) is one of the major plant growth-limiting nutrients although it is abundant in soils in both inorganic and organic forms [7,8]. P is vital for plant cell division, growth, root lengthening, seed and fruit development, and early ripening as well, it is a part of several compounds including oils and amino acids in plants [1]. P-supplementation can enhance plant growth by increasing the efficiency of biological nitrogen fixation, enhancing the availability of macronutrients such as N, P, K, Ca and Mg [5,9]. It has been reported that phosphorus application influences the content of others nutrients in leaves and seeds e.g. Zn, Mn, Fe, B, Mo etc. [10]. Microorganisms such as Bradyrhizobium inoculants may have effects on the chemistry of nutrients in soils by enhancing nutrients uptake by plants. For example, [5] reported improved uptake of macronutrients following inoculation with efficient strains of Rhizobium. Studies on the influence of P and inoculation for the successive uptake of macronutrients in different plant organs are scarce in Africa. This study was conducted with the objective to evaluate the effects of B. japonicum inoculation and phosphorus supplementation on the uptake of macronutrients in soybean.
2. Materials and Methods
2.1. Narrative of Site Location
Glasshouse and field experiments were conducted at Selian Agricultural Research Institute (SARI) and Tanzania Coffee Research Institute (TaCRI) at Northern part of Tanzania. The coordinates of SARI lies at 3˚21'50.08''S and at 36˚38'06.29''E at an elevation of 1390 masl with mean annual rainfall of 870 mm. The mean maximum temperature ranges from 22˚C to 28˚C whiles the mean minimum temperature ranges from 12˚C to 15˚C. TaCRILyamungu laid at 3˚13'59.59''S and 37˚14'54''E, at an elevation of 1268 masl with means annual rainfall of 1200 mm. The mean maximum temperature is about 21.7˚C and the mean minimum temperature is about 13.6˚C with relative humidity of about 94%. The field experiment was carried out during the long rain season while the glasshouse was conducted under controlled conditions during 2013 cropping season. The physical and chemical characteristic of the soil were shown in Table 1.
2.2. Experimental Design
Both the glasshouse and field experiments had 2 levels of rhizobia inoculation (with and without rhizobia) and 4 levels of P-supplementation (0, 20, 40, and 80 kg P∙ha−1). The experimental design pursued a split plot with 4 replications per treatment. The soybean seeds were seeded at a spacing of 50 cm by 20 cm while the plot size measured at 4 m by 3 m. The plant population was 200,000 plants per hectare. The Bradyrhizobium japonicum strain

Table 1. Chemical & physical characteristics of the soil.
USDA 110 (Batch number 23011302, S) was obtained from MEA Company Nairobi-Kenya, sold under license from the University of Nairobi. Soybean seed (Soya 2 variety) was obtained from Uyole Agricultural Research Institute-Mbeya-Tanzania. Before sowing, soybean seed were thoroughly mixed with B. japonicum inoculants to supply (109 cells/g seed), following procedures stipulated by products manufacturer. To avoid contamination, the un inoculated seed was planted first followed with the inoculated seed. Three seed were planted and thinned to two plants after full plant establishment. Weeding and other agronomic practices were done manually using hoe at different growth stage of the soybean plant.
2.3. Plant Harvest and Sample Preparation
Before flowering, plant samples from glasshouse and field experiments were collected for plant nutrients analysis. For field experiment, 10 plants were randomly sampled from the middle rows of each plot. The same procedure used for the glasshouse, where two plants from each pot were sampled. The sampled plants were carefully dug out with their entire root system, washed and separated into roots, shoots and pods. The plant parts were oven dried at 60˚C for 48 hours, ground into a fine powder (2 mm sieve) for N, P, K Ca, Mg and Na analysis.
2.4. Determination of Nutrients in Soybean Plant Organs
Total N was determined by the micro-Kjeldahl method [11]. Phosphorus was determined by the molybdenum blue method as described by [12]. Ca, Mg, K and Na concentrations in plant extracts were determined by method described by the [13].
2.5. Statistical Analysis
A 2-way ANOVA was used to compare between this difference treatments to analyze the nutrients in soybean plant organs. The analysis was done using the STATISTICA software program 2013. Fisher’s least significant difference was used to compare treatment means at p = 0.05 [14].
3. Results
3.1. Effects of B. japonicum Inoculation on Macronutrient Uptake in Roots, Shoots, Pods and the Whole Plant of G. max L.
B. japonicum inoculation of soybean plants significantly increased root uptake of some of the macronutrient measured in field and glasshouse relative to the control treatments (Table 2). In the glasshouse, the uptakes of all macronutrients in roots were numerically increased with inoculation. However, significant root uptake was observed only in N. For the field experiment, the root uptake of N, P, K, Ca and Mg were significantly increased with B. japonicum compared with the un-inoculated control (Table 2). With shoots, of B. japonicum significantly increased the uptake of N and P in the glasshouse, while in the field experiment the uptake of N, P, K, Ca and Mg were significantly increased in shoots relative to the control treatment (Table 3). In the pods, inoculation significantly increased the uptake of N and P in the glasshouse experiments, while for field experiment, significant uptake were observed of N, P, K Ca and Mg (Table 4). Inoculation significantly increased the whole plant uptake of N, P and Ca for glasshouse experiment, whereas the uptake of N, P, K, Ca and Mg were significantly increased with inoculation in field experiment (Table 5).
3.2. Effects of P Supplementation on Macronutrient Uptake in Roots, Shoots, Pods and the Whole Plants of G. max L.
By supplying P the root uptake of N and Na were significantly increased in glasshouse experiment while in field experiment the root uptake of N, K and Na were significantly increased with P supplementation relative to the control (Table 2). In the glasshouse experiment, P supplementation significantly increases the uptake of N, P and Mg relative to the control treatment (Table 3). In pods, P supplementation significantly increased the N, P and Mg content in glasshouse experiment while in field experiments; P supply significantly increased the uptake of N, Ca and Na respectively relative to the control (Table 4). In whole plants, P supply significantly increase the uptake of N, P, Ca, Mg and Na except K in the glasshouse experiment, while in field experiment, P supply significantly increase the uptake of N, P and Na relative to the control treatment (Table 5).
3.3. Interactive Effects of B. japonicum Inoculation and P Supplementation on Macronutrient Uptake in Soybean Roots
Significant interaction between B. japonicum and P supplementation was observed in root N concentration in the field experiment (Figure 1). Combining B. japonicum and P increased the N in roots with the higher levels obtained at the highest P level of 80 kg P∙ha−1. Low N content was observed in un-inoculated treatments (Figure 1).
4. Discussion
B. japonicum inoculation significantly increased the uptake of macronutrients in the roots, shoots, pods and the whole plant of soybean grown in glasshouse and in field experiment (Tables 2-5). There was a significant increase in N, P, K, Ca and Mg in all plant organs measured in the field experiment and few in the glasshouse study. The positive results especially in field experiments might be influenced by enough soil volume thus increased root length which facilitated the plants to capture nutrients in the nearby plant roots and allow for more nutrients uptake. Furthermore, production of plant growth hormones and secretion of chemical substances by rhizobia bacteria might have stimulated plant growth and ultimately increased the uptake of macronutrients in plant tissues as suggested in different studies [15-17]. It has also been reported that the rhizosphere tends to be modified by the released dead cells of rhizobial inoculants. The released dead cells may contain macroelements or biomolecules that can solubilise unavailable soil nutrients [18-22]. B. japonicum inoculation may have in-
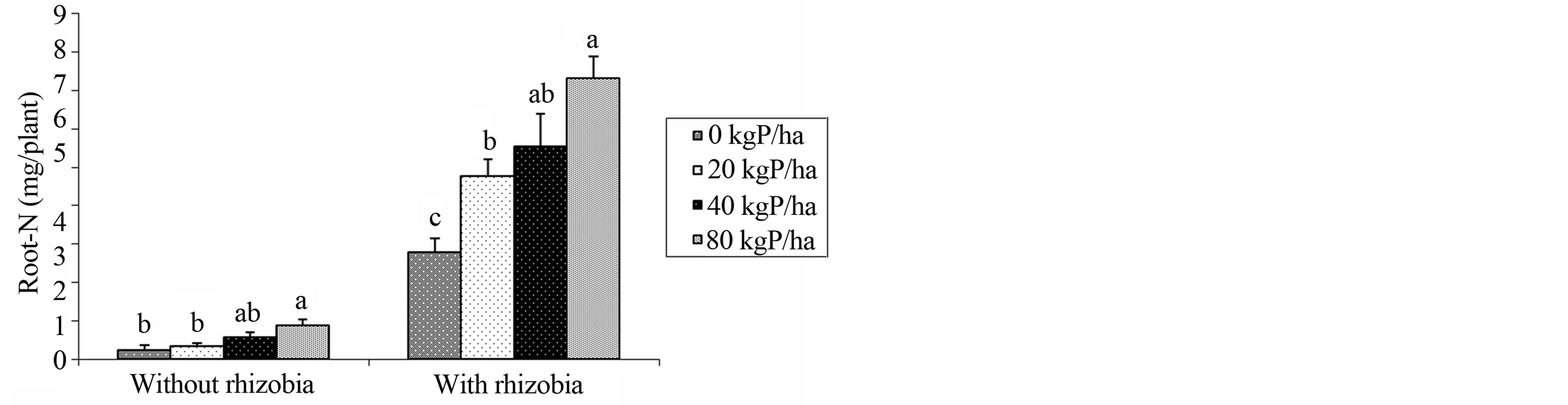
Figure 1. Interactive effects of B. japonicun and phosphorus on N uptake in roots. Bar(s) followed by dissimilar letters are significantly different by Fisher Least significant difference (LSD) test at p = 0.05.

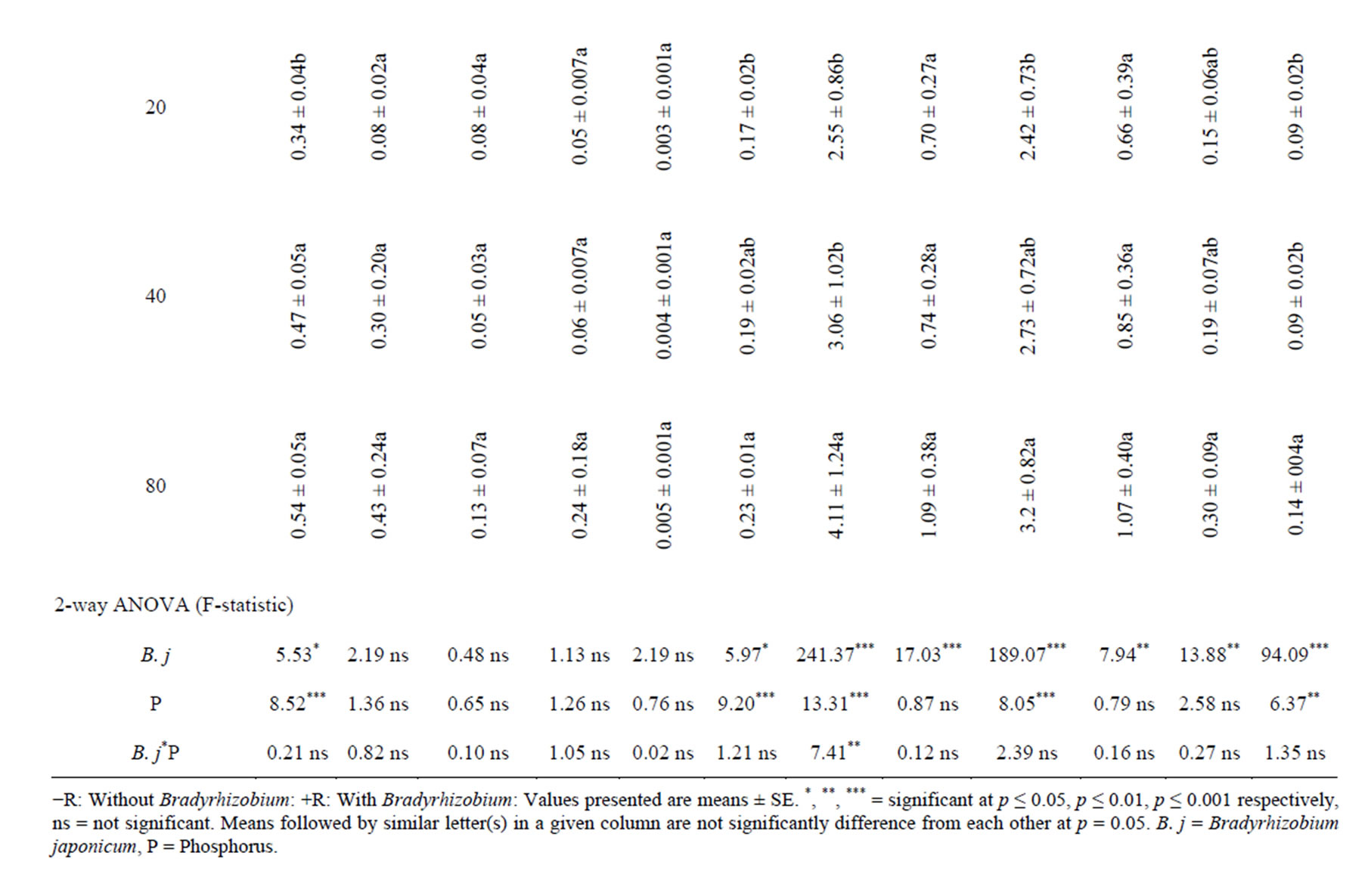
Table 2. Effect of B. japonicum, and P supplementation on macronutrient uptake in roots of G. max L.

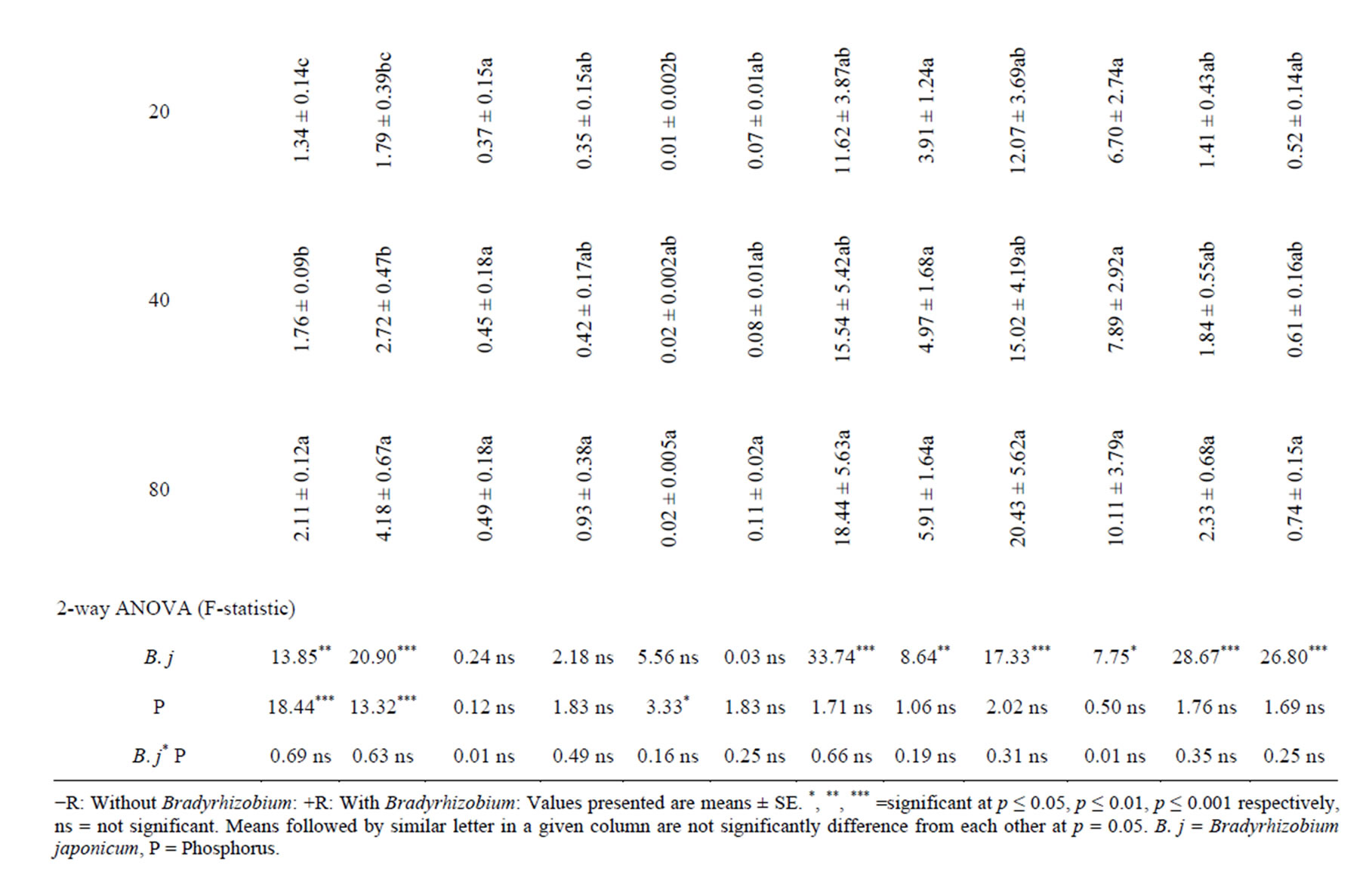
Table 3. Effect of B. japonicum, and P supplementation on the macronutrient uptake in shoots of G. max L.


Table 4. Effect of B. japonicum, and P supplementation on the macronutrient uptake in pods of G. max L.
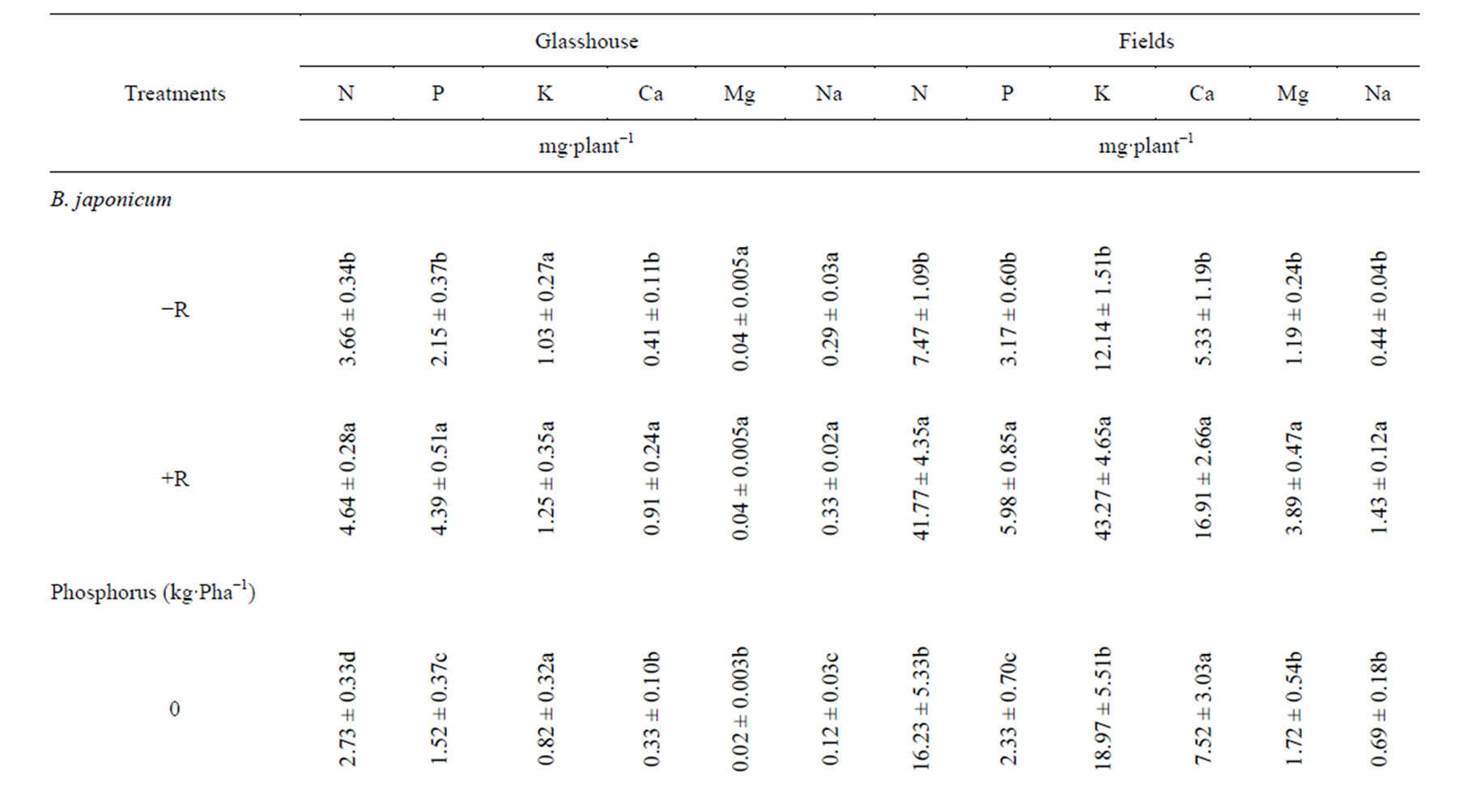

Table 5. Effect of B. japonicum, and P supplementation on the macronutrient uptake in the whole plant of G. max L.
creased their biological activity associated with roots of the host plant and the microorganism, thus increasing the decomposition of organic matter in the soil. Such root residue decomposition may have enhanced root exudation and hence increasing the amount of available nutrients in the soil [23-25]. The potential of some rhizobia bacteria to stimulate nutrient uptake in different legumes tissues was also reported by [5,26,27].
In the present study, P supply leads to the significantly increase in the uptake of some macronutrient such as N, P, K, Ca and Mg both in glasshouse and field experiment of the G. max plant organs respectively (Tables 2-5). The role of phosphorus in plants is to facilitate root growth [28]. For this reason, root growth in this study was enhanced by supplying P and hence enabling the plants to explore for more water and nutrients such as N, P, K, Ca, Mg from the soil solution and ultimately making them available to different plant organs as observed in the present study.
The data obtained from our study showed significant interaction in the uptake of N in roots in the field experiment (Figure 1). Combining B. japonicum and P increased the uptake of N in roots whereby at the highest P levels (80 kg P∙ha−1) the uptake of N in roots was superior to all other treatments. Similar to our study, Hussain and Ali 2012, [29] report that the combination of phosphorus and inoculation showed maximum positive effect on nitrogen uptake in mungbean.
In conclusion, B. japonicum and phosphorus supplementation had positive impacts in increasing nutrient uptake in G. max in Africa. These inputs in one way enhanced the availability of N, P, K, Ca and Mg. This was advantageous to cropping systems involving G. max as they require greater amounts of these nutrients for normal plant growth and yield.
Acknowledgements
This study was supported by Tanzania Commission for Science and Technology (COSTECH) and Nelson Mandela African Institution of Science and Technology (NMAIST)-Tanzania. The Tanzania Coffee Research Institute and Selian Agricultural Research Institute are acknowledged for providing the study sites.
NOTES