A Plasmid vector encoding functional human keratinocyte growth factor gene in vitro—Functional human KGF gene expression in vitro ()
1. INTRODUCTION
It has become clear that type II alveolar epithelial cell (AEC II) proliferation is not only important in physiological processes of fetal and neonatal lung development in association with somatic growth, mechanical stretching, physical, chemical and biological stimulation, but also plays a pivotal role in pathological conditions, such as lung inflammation, injury and reparation due to hyperoxia and hypoxia, barotrauma and volotrauma, radiation, and other natural or iatrogenic hazards [1-3]. Growth factors that are mitogenic for adult rat, rabbit, or human AEC II in vitro include epidermal growth factor, transforming growth factor-α (TGF-α), insulin, and acidic and basic fibroblast growth factor (FGF).
Keratinocyte growth factor (KGF; also known as FGF-7), a member of the FGF family, is a potent mitogen specific for epithelial cells [4]. Unlike other members of this family, KGF is produced by cells of mesenchymal origin of several stromal fibroblast lines derived from embryonic, neonatal, and adult sources [5,6]. In these tissues, KGF functions in mesenchymal induction of epithelial growth as a stromal-derived paracrine regulator of epithelial proliferation. It is important in fetal pulmonary growth and differentiation [7]. A protective role of KGF to block apoptosis and to prevent oxidative damage has also been suggested for AECs, hepatocytes, and keratinocytes [1,2,8,9]. KGF induces AEC II proliferation in vitro and in vivo [3,10,11]. Furthermore, lungs of transgenic mice expressing a dominant negative for the KGF receptor (FGF receptor 2-IIIb) under control of the surfactant protein C promoter have grossly abnormal lung development, with only two primordial epithelial tubes and no branching morphogenesis [12]. Transgenic mice overexpressing KGF exhibit lethal papillary cystadenoma, with marked enlargement of the bronchial air spaces [13].
In recent years, the therapeutic potential of these growth factors in lung disease, especially in lung injury and reparation, has yet to be fully explored [3]. KGF has been best studied and has impressive protective effects against a wide variety of injurious stimuli when given as a pretreatment in animal models. Whether this protective effect could translate into a treatment effect in humans with acute lung injury (ALI) needs to be investigated [3,14]. Regardless of their therapeutic potential, studies of the unique roles played by these growth factors in the pathogenesis and the resolution of ALI and other lung diseases will continue to enhance our understanding of the complex pathophysiology of inflammatory injury and repair in the lung. Recombinant eukaryotic expression vector is one of the most popular and promising tools for gene delivery [15]. But how to construct a proper recombinant vector carrying the interested gene has become the bottle neck which restricts the application of the vector in clinical gene therapy. In the present study we sought to construct a eukaryotic expression vector containing hKGF gene, which was capable of introducing exogenous gene in vitro, so as to enable further experimental foundation for ALI study.
2. MATERIALS AND METHODS
2.1. Bacterial Strains, Plasmids, and Cultivation
For preparation of total DNA, human embryo lung fibroblast was cultivated. E. coli DH5α and an expression vector pMD18-T Vector (Takara Bio, Dalian, China) were used for cloning and expression studies. E. coli DH5αand an expression vector pEGFP-C2 used for eukaryotic expression. E. coli strain were cultivated in Luria-Bertani (LB) medium supplemented with antibiotics (100 μg/mL ampicillin and/or 35 μg/mL chloramphenicol) when they were required. The recombinant plasmid pEGFP-C2-hKGF ncludes hKGF genes from human embryo lung fibroblast in pEGFP-C2. The nucleotide sequence of hKGF is available from GenBank under Accession No. M60828.
2.2. Human Embryo Lung Fibroblast Cell Culture
human embryo lung fibroblast was cultivated were obtained from cell Bank and seed into 12-well plates and cultured in growth medium DMEM supplemented with 10% heat-inactivated fetal bovine serum (FBS), 100 U/mL penicillin, and 100 μg/mL streptomycin in a humidified atmosphere containing 5% CO2 and 95% air at 37℃. After six days of differentiation, human embryo lung fibroblast were grown in monolayers until they reached a confluency of 70-80%.
2.3. RNA Isolation and CDNA Synthesis
Total RNA was isolated from monolayers of human embryo lung fibroblast using the guanidine isothiocyanate based TRIzol solution (GIBCO-BRL, Burlington, ON, Canada) according to the manufacturer's specifications. The RNA samples were resuspended in 100% formamide and quantified spectrophotometrically at 260 nm. All RNA isolates had an OD260:OD280 between 1.8 and 2.0, indicating clean RNA isolates. The RNA quality was also checked by 1.0% agarose gel electrophoresis, stained with 1 ug/ml ethidium bromide. Oligo-(dT)18 (Invitrogen, Burlington, ON) was used as primer in the first step of cDNA synthesis. Total RNA (5 μg) was combined with 0.5 μg oligo-dT, 200 μM dNTPs and H2O and preheated at 65℃ for 2 min to denature secondary structures. The mixture was then cooled rapidly to 20℃ and then 10 μL 5 × RT Buffer, 10 mM DTT and 200 U MMLV Reverse Transcriptase (Sigma-Aldrich, Oakville, ON, Canada) was added for a total volume of 50 μl. The reverse transcription mix was incubated at 37℃ for 90 min. then stopped by heating at 95℃ for 5 min. The cDNA stock was stored at –20℃. The yield of cDNA was measured according to the PCR signal generated from the internal standard house-keeping gene β-actin amplified from 18 to 24 cycles starting with 0.1 μL of the cDNA solution. The volume of each cDNA pool was adjusted to give the same exponential phase PCR signal strength for β-actin after 20 cycles.
2.4. Amplification of the HKGF Gene
The hKGF gene (GenBank Accession No. M60828) was amplified by PCR from the cDNA solution as described above, using two oligonucleotide primers P1 (5’-CCT AGA TCT GCC ACC ATG CAC AAA TGG ATA CTG AC-3’, the BglII restriction site is underlined and the ATG initiation codon is indicated by boldface type) and P2 (5’-CCT CTC GAG TTA AGT TAT TGC CAT AGG AAG-3’, the XhoI restriction site is underlined and the TTA termination codon is indicated by boldface type), with the aid of the computer program Primer Premier 5.0. The sense primer is homologous to nucleotides 1 to 20 in the hKGF open reading frame and contains a consensus (GCCACC) Kozak sequence immediately upstream of the initiation codon. In addition, a BglII site was inserted at the 5’-terminus to facilitate cloning. The antisense primer is homologous to nucleotides 565 to 585 and contains an XhoI recognition sequence at its 5’ terminus. These primers allowed amplification of a 609-bp cDNA fragment16. The PCR process involved at 95℃ for 2 min, and then 35 cycles of 95℃ for 30 s, 60℃ for 1 min, 72℃ for 1 min. at the last cycle, PCR products were extended by keeping the temperature at 72℃ for 10 min. Then 10 µl of each PCR product was electrophoresed on 1% agarose gel containing 0.5 µg/mL EB, and PCR products were then purified from agarose gel according to the protocol of DNA purification kit. The resulting PCR fragment was blunted by T4 DNA polymerase, phosphorylated by T4 polynucleotide linase, and ligated to the ScaI-digested and phosphorylated pMD18-T Vector by T4 DNA ligase. The resulting recombinant plasmid was mixed with E. coli DH5α cells (1:19), on ice for 30 min, 42℃ 2 min, then added 2 mL LB, ice for 2 min, 37℃ 80 min, and added to X-gal 40 μL, IPTG 20 μL, 37℃ overnight. Transformed cells were spread onto LB medium agar plates for blue-white blot screening. The positive clones were selected from the transfected E. coli DH5α cells. Blue colonies due to indigo production on LB agar plates were selected as candidates for cells carrying hKGF gene from E. coli DH5α cells. The recombinant plasmid was isolated from E. coli DH5α cells using Plasmid Extract Kit. The insert was digested and sequenced to ensure that the correct construction had been obtained.
2.5. Construction of an Expression Vector
The amplified recombinant plasmid was digested with endonucleases BglII and XhoI then followed with electrophoresis. The amplified hKGF fragment was retrieved using PCR Fragment Recovery Kit (Takara, Bio, Dalian, China) according to manufacturer’s guide. Recovery hKGF cDNA was ligated to pEGFP-C2 digested by BglII and XhoI. Briefly, 1 μL pEGFP-C2, 1 μL cDNA fragment and 5 μL Universal Buffer were mixed at 4 ℃ overnight. The pEGFP-C2 plasmid inserted with correct hKGF gene was simultaneously introduced into E. coli DH5α cells. The recombinant plasmid was isolated, digested and sequenced to get correct construction plasmid of pEGFP-C2-hKGF.
2.6. S-D Rat AECII Cells Isolation
Alveolar epithelial cells were isolated from pathogen-free male Sprague-Dawley rats (180-200 g) as previously described [10]. The cell pellet (70% purity, > 95% viability, 8-10 × 106 cells/rat) was plated at a density of 7-10 × 105 cells/cm2 in six-well culture dishes. Culture medium consisted of DMEM containing 25 mM D-glucose, 10 mM HEPES, 23.8 mM NaHCO3, 2 mM L-glutamine, 10% FCS, 50 U/mL penicillin, 50 µg/mL streptomycin, and 10 µg/mL gentamicin incubated in a 5% CO2-95% air atmosphere. Culture medium was changed 24 h after isolation and then on alternate days.
2.7. Cell Transfection, Cell Count
Cells were seeded in a 6-well plate with 2 × 105 cells per well 24 h prior to transfection when they were cultured to a confluency of about 90%. Cell transfection was performed according to the manufacturers instructions. Briefly, a transfection mixture was prepared by adding 6 mg of plasmid DNA and 20 mL lipofectame (GIBCO BRL) to 500 ml serum-free RPMI1640. After incubated at room temperature for 20 min, the transfection mixture was added to the cells to be cultivated for 4 h at 37℃ when the media containing the transfection mixture was exchanged for growth medium. After transfection cells were observed and photographed by fluorescence microscope and counted at 24, 48, 72, and 96 h.
2.8. Expression Analysis by Immunohistochemistry
The gene transferred AECII cells were washed and fixed with 4% paraformaldehyde then incubated with the 1:200 dilution of rabbit anti human primary antibody to KGF (Santa Cruz Biotechnology, Santa Cruz, CA) or control IgG (1:1000) overnight at 4℃. The tissue sections were washed in PBS, then incubated with a 1:300 dilution of biotinylated secondary sheep anti-rabbit or goat anti-rabbit IgG. After washing with PBS, tissue sections were incubated with an avidin-biotin complex and developed in 0.075% (w:v) 3, 3 diaminobenzidine (DAB). After lightly counterstaining with haematoxylin, the sections were dehydrated. The intensity of immunostaining around the stent struts was scored as follows: 0, no staining; 1) minor staining only; 2) moderate staining; and 3) heavy staining. Scoring was performed on every third strut in each vessel beginning with the strut closest to the top of the slide by an investigator blinded to the treatment allocation.
2.9. Akt Kinase Assay
Akt kinase assay was performed using Akt Kinase Assay kit (Cell Signaling Technology, Beverly, MA), following the manufacturer's instructions. In brief, soluble extract of lung tissue was prepared by using Triton X-100containing extraction buffer and used for immunoprecipitation of Akt with anti-Akt antibody conjugated to agarose beads. The immunoprecipitated Akt was incubated with glycogen synthase kinase (GSK)-3 protein in kinase assay buffer containing ATP. After centrifugation, an aliquot of the supernatant was removed and tested for the presence of phospho-GSK-3 by western blotting using anti–phospho-GSK-3α/ß (Ser 21/9) antibody. Equal amounts of soluble extracts were immunoprecipitated with anti-Akt Ab, and The Akt bound to the agarose beads was released by boiling in SDS-sample buffer and subjected to western blot analysis.
3. RESULTS
3.1. The HKGF Gene Fragment Amplified by PCR from Human Embryo Lung Fibroblast
To clone the full sequence gene encoding human KGF protein for functional expression, standardized RT-PCR assay was performed for cloning the gene from the cDNA of human embryo lung fibroblast. Total cellular RNA was purified and RT-PCR was performed using primers based on the published hKGF cDNA sequence, as described in methods. Conditions and cycle number were optimized for primer. This yielded target fragments of hKGF gene with expected 609 bp in length that was subsequently TA-cloned into a pMD18-T vector (Figure 1).
3.2. Enzyme Digestion Analysis of the Recombinant Plasmid
We next analyzed the sequence of the fragment of PCR-amplified hKGF cDNA (609 bp). The recombinant

Figure 1. Generation of hKGF cDNA amplified from human embryo lung fibroblast. M: marker (500bp DNA ladder), 1: Lane 1 shows the PCR product (arrow shown), corresponding to the hKGF cDNA (609 bp). Samples were run on a 3% agarose gel with a DNA ladder.
plasmid inserted hKGF cDNA were both doubly digested with PST1/ECOR1 (Figure 2). A 601 bp fragment was released from pMD18-T. This feature of exogenous gene fragment introduced argues in favor of a successful clone.
3.3. Nucleotide Sequence Analysis
Further analysis of the nucleotide sequences of hKGF gene in pMD18-T revealed that the sequences of the cloned gene was completely coincidence with the published hKGF sequences [16].
3.4. Insertion of HKGF Gene into PEGFP-C2 Plasmid
To construct eukaryotic expression vector, PCR products were subcloned into a pEGFP-C2 vector. The construction strategies of recombinant exogenous hKGF gene and eukaryotic expression plasmid pEGFP-C2-hKGF are described elsewhere. Recombinant gene hKGF was 609 bp with PST1/ECOR1 restriction site on each side, the size of pEGFP-C2-hKGF was 5.3 kb.
3.5. Identification of Eukaryotic Expression Vector AEGFP-C2-HKGF
The eukaryotic expression vector pEGFP-C2-hKGF was identified by restriction endonucleases cut with PST1/ECOR1 and electrophoresis generating a 601 bp fragment according to the result from GeneBank (Figure 3). By nucleotide sequencing, coinciding completely with the sequence from GeneBank. There were no endonuclease cut sites of PST1/ECOR1 by zymogram analysis.

Figure 2. The recombinant plasmid was doubly-digested with PST1/ECOR1. M: marker (1 kb DNA ladder), 1: the product of PCR doubly digested with PST1/ECOR1, yielding expected 601 bp and 2692bp cDNA fragments.

Figure 3. Expressing vector pEGFP-C2-hKGF was digested by PST1/ECOR1. M: Marker (1 kb DNA ladder); 1. 601 bp and 4.7 kb DNA fragments were appeared as expected.
3.6. Expression of the Target HKGF Protein by Immunohistochemistry
AECII cells from S-D rat transfected by either pEGFPC2-hKGF (the experimental group) or pEGFP-C2 (the control group) for 24, 48, 72 h were fixed, paraffin embedded, and then immunostained with an antibody raised against KGF and KGFR, which cross-reacts with porcine KGF and KGFR. We found scattered AECII staining at 24 h group (mean score 0.5 ± 0.5). Expression appeared strong at 48 h (mean score 1.8 ± 0.8) and appeared to peak at 72 h (mean score 2.1 ± 0.8) (Figure 3(b)). But, there was no statistical significant between 48 h and 72 h groups (Figure 4, P > 0.05). KGF expression was undetectable at 24 h and 48 h control groups. The results of fluorescence expression were similar to that of immunohistochemistry (Figure 5).
3.7. Effect of HKGF on AECII Cell Growth
Cell growth was measured by cell count. The cell numbers were 2.8 × 106, 3.6 × 106, and 5.0 × 106 in the experimental group and 2.6 × 106, 3.2 × 106, and 3.9 × 106 in the control group at 24, 48, and 72 h respectively. The growth of AECII was stimulated by KGF in a timedependent manner (Figure 6, P < 0.05).
3.8. KGF-Induced Activation of Akt in Vivo
We next have investigated whether high KGF expression follow up a dramatic increase in Akt activation as evaluated by Akt kinase assay. We have shown that expression of constitutively active Akt appeared strong transfected by eukaryotic expression vector pEGFP-C2-hKGF (Figure 7). Thus, one possible explanation for KGF-mediated effect is associated with activation of the Akt signaling axis.
4. DISCUSSION
This Eukaryotic expression plasmid vector expressing KGF in our study was designed for two purposes. The first was to study the underling mechanism of KGF such as producing extensive type II cell hyperplasia. It is much more difficult to find this extensive type II cell hyperplasia in rodent lungs with a variety of injuries, including bleomycin. Therefore in rodents, it has been difficult to investigate the biologic properties of hyperplastic type II cells and their ability to alter the inflammatory and fibroproliferative response. Type II cells can augment fibrotic reaction by producing PDGF, TGF-α, and TGF-β [17-20]. However, type II cells also inhibit fibroblast proliferation [21]. Exogenous KGF should make it possible to study the biologic properties of hyperplastic type II cells. The second purpose was to have a means of delivering KGF for a sustained period in a local area for the treatment of lung injury. Currently, because of the inflammation induced by adenoviral vectors, therapy of acute lung injury may not be possible. Another approach is to decrease the host response by using plasmid vector.
Over the past 10 yr, it has become increasingly clear that KGF play important roles in both the normal and the injured lung and ultimately may have therapeutic potential as a targeted therapy to facilitate lung epithelial repair in lung disease. KGF has been shown to increase the expression of SP-A and SP-D in cultured alveolar type II cells in vitro [22,23]. Administration of KGF has been
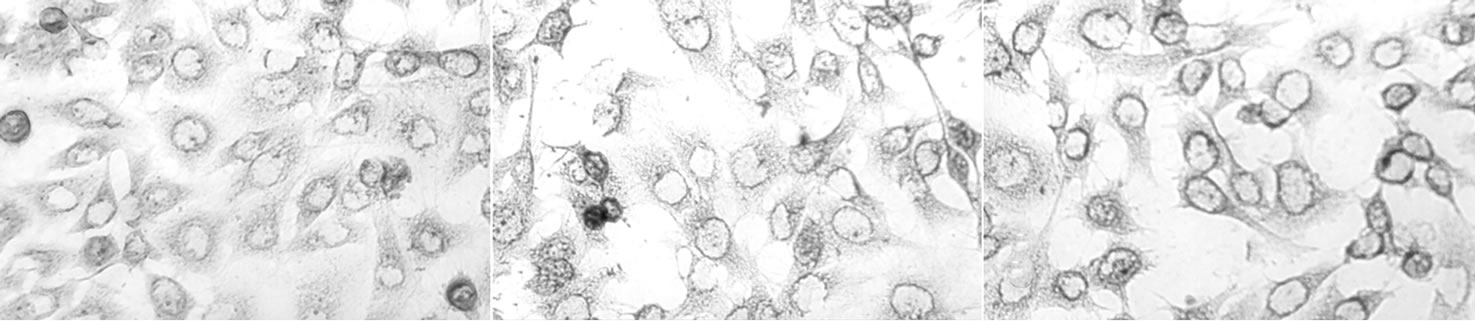 (a) (b) (c)
(a) (b) (c)
Figure 4. Immunolocalization of KGF in S-D AECII cells. Positively stained cells appear brown color. Positive immunoreactivity to KGF is present in surrounding cytomembrane and is weak in the cytoplasm. The control group data did not show. Note the strong cytomembrane staining in S-D AECII cells 72 h transfection with hKGF vector (c), whereas moderate in S-D AECII cells 48 h transfection with hKGF vector (b) and weak in S-D AECII cells 24 h transfection with hKGF vector (a). Original magnification × 200.
 (a) (b) (c)
(a) (b) (c)
Figure 5. Fluorescence expression in S-DAECII cells. Positively stained is weak at 12 h (a), and strong at 48 h (b), peak at 72 h (c). The control group data did not show. Original magnification × 200.

Figure 6. Cell count of AECII cells transfected by eukaryotic expression vector pEGFP-C2-hKGF in different time (× 106). Each point is mean ± SE from 6 observations from a representative experiment. Where no error bars are shown, they are included in the point. Similar results were obtained in 2 further experiments.

Figure 7. Activity of Akt kinase in the AECII by eukaryotic expression vector pEGFP-C2-hKGF. Akt kinase activity of AECII transfected with eukaryotic expression vector pEGFPC2-hKGF for 72 h (b) was detected in a high level as compared with normal AECII (a) and vector control (c). The blot was stripped and probed with anti-Akt Ab as described in Materials and Methods and shown equal amounts of Akt in immunoprecipitates. Three observations were included per condition and data shown are representative of two independent experiments.
shown to protect the lung from a variety of insults including oxygen, radiation and chemotherapy. In bleomycin-induced lung injury, KGF was shown to decrease lung edema.
However, the KGF has had to be given before the injury in order to be effective. A more sustained delivery of KGF or expression of KGF might allow for successful post-treatment therapy. Gene transfer into specific tissues or cell types is a key technique in the development of gene delivery strategies study, gene function research and gene therapy. Adenoviral vectors can efficiently transfer foreign genes into lung tissue in vivo [24,25]. However, adenovirus-mediated gene expression is transient and may be associated with a significant inflammatory response, depending on the viral dose [26]. Unfortunately, this inflammatory response limits the use of these vectors in treating acute lung injury. The host response is mainly to the virus.
Eukaryotic expression plasmid vector capable of directing the expression of exogenous gene has been found to be potentially valuable for the study of specific gene function and for gene therapy [27-29]. We described the establishment of a eukaryotic expression system, which allows us to express hKGF in S-D rat AECII. The versatility of this system is evident in our ability to express hKGF. Because our objective in this study was to study the effects of KGF, it was important to develop this system. The results of immunohistochemistry performed in this study demonstrate that the constructed eukaryotic expression system pEGFP-C2-hKGF efficiently produces hKGF gene. And cell count result suggested that hKGF could induce S-D rat AECII cell proliferation.
As plasmid vector gene transfer is one of the most reliable and convenient methods for introducing genes into almost all types of mammalian cells and for expressing the genes at high levels since many cells receive multiple copies of the recombinant genome [30-32]. Gene delivery using eukaryotic expression plasmid vector that propagates in cultured cells might be an effective way for research [33,34]. In order to increase the effect of putative gene therapy, in this study, primers containing specific enzyme-cutting sites were designed to amplify the 0.6 kb hKGF from human genome, and the 0.6 kb sequence was cloned into the eukaryotic plasmid. The hKGF gene, cloned from human embryo lung fibroblast, showed high homologies of nucleotide and putative amino acid sequences compared with the published corresponding sequences [35,16].
It was shown that there are several different signal transduction pathways to be stimulated by growth factors (36) and through Akt signaling to transmit induced apoptosis signaling to link life and death decisions (37). our data support the idea that high levels of KGF positively regulate Akt during lung development. KGFinduced Akt activation may play an important role in inhibiting lung alveolar cell death thereby preserving the lung architecture and function during oxidative stress (38). At present, we have no simple explanation for how KGF modifies the response of Akt signal. Future insight into the relationships between KGF and Akt in vivo will come from functional and biochemical studies on the pathway of the two ligands, and from the study of both KGF-deficient and KGF, Akt double null mutant mice.
In summary, we succeeded in constructing the hKGF eukaryotic expression vector pEGFP-C2-hKGF acting as a mediating mesenchymal-epithelial interactions, and studied its bioactivity of induction type II cell proliferation. But the molecular mechanisms of pEGFP-C2hKGF mediated proliferation of AECII cells still need to be established.
5. ACKNOWLEDGEMENTS
We thank Prof. Wang Fu-Liang for providing technical assistance and insightful discussions during the preparation of the manuscript. This work was supported by National Scentific Foundation of China (No: 30030781).