Electroencephalographic Activity Associated to Investment Decisions: Gender Differences ()
1. Introduction
Literature in finance and neurosciences describes that, on average, women are more risk averse than men when making financial decision investment (e.g., [1] ). Becker et al. [2] defend that males are more likely than females to engage in risky behaviors and that gender differences are due, at least in part, to sex differences in the organization of the neural systems responsible for motivation. Zaidi [3] wrote that “men and women appear to use different parts of the brain to encode memories, sense emotions, recognize faces, solve certain problems and make decisions”. He also pointed out that “women use a variety of parts of the brain when they do a simple task∙∙∙ Male brains separate language in left, and emotions in the right, while female’s emotions are in both hemispheres”. Haier et al. [4] proposed that “… Such finds suggest that human evolution has created two different brains (male and female ) designed for equally intelligence”, a fact also confirmed by Rocha et al. [5] .
Lee et al. [6] analyzed gender effects on the process of risk-taking and found a stronger activation in the right insula and bilateral orbitofrontal cortex in female than the male participants while they were performing in the Risky-Gains task. When taking the same level of risk, relative to men, women tend to engage in more neural processing involving the insula and the bilateral orbitofrontal cortex to update and valuate possible uncertainty associated with risk-taking decision making. Burgdorfa and Pankseppa [7] , and Xue et al. [8] complement this information describing that insula is activated during the decision process, using past experiences to make future decisions.
Based on Iowa Card Task, Bolla et al. [9] found that men and women activated different parts of the brain when solving the same decision-making task. Using the same test, Overman [10] found that males and females had a different response pattern. Females tended to choose cards associated with immediate wins and males tended to choose cards related with long-term outcome meaning that women preferred investments that produced short term outcomes. Lighthall [11] complemented this investigation describing that when they manipulated the stress level, the impact of stress on reward-related decision processing differed depending on gender.
Vieito et al. [12] used EEG technology to study financial decision in a simulated traded decision game. In this study, two groups’ volunteers (G1 and G2) made decisions about buying and selling of holding stocks of 7 different companies in two market conditions (M2 and M2) differing in stock volatility, while their brain activity was recorded. Market M1 is a bull market with prices steadily increasing and having low volatility. Market M2 has high volatility with some stocks experiencing losses while some others experience slow gains. Group G1 traded first (trading session S1) on M1 and later on M2 (trading session S2). In contrast, group G2 traded first (S1) on M2 and latter (S2) on M1. Each experimental group was composed by 10 males and 10 females from a business school with no previous experience on the stock market. In this way, previous knowledge influence upon the experiment was minimized because volunteers attended the same financial courses and had no trading experience. These authors showed that in such conditions, experimental groups G1 and G2 learned to successfully play the game because their portfolio value plush cash (total amount money earned on selling minus total amount spent on buying) was above the market value of their 1400 stocks. Using Principal Component Analysis, they showed that brain activity during trading differed for G1 and G2.
Electroencephalogram uses electrodes placed over the skull to record the electrical fields 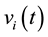 generated by ionic currents triggered by cortical activity supporting a cognitive task. sLORETA (Low Resolution Tomography) uses measurements of scalp
generated by ionic currents triggered by cortical activity supporting a cognitive task. sLORETA (Low Resolution Tomography) uses measurements of scalp 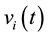 to find the 3D distribution of the activated cortical areas, with exact zero error localization to point-test sources [13] . This technique has been widely used in studies covering very different aspects of brain physiology, and it allows identification of the sets of cortical neurons
to find the 3D distribution of the activated cortical areas, with exact zero error localization to point-test sources [13] . This technique has been widely used in studies covering very different aspects of brain physiology, and it allows identification of the sets of cortical neurons 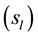 activated during a cognitive task processing.
activated during a cognitive task processing.
The purpose of the present paper is to apply sLORETA to disclose the sources of the EEG recorded 2 seconds before trading decision making in order to test the hypothesis, supported by the literature discussed above, that male and female enroll different neural circuits to make equally successful financial decisions trading on both M2 and M2.
2. Methods
Experimental Design
The investigation was done based on a sample composed by 40 undergraduate students of the School of Business Studies from Polytechnic Institute of Viana do Castelo, Portugal. The sample is composed by 20 women and 20 men from age 20 to 45 years.
Trading took place in two distinct market conditions: Market 1 (M1) with steadily increasing prices and Market 2 (M2) with randomly changing prices and high volatility.
Daily stock prices were collected from trading in FMBovespa (the Brazilian Bourse) from January 4, 2010 to December 30, 2012. Collected stock prices 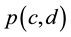 were divided by the stock prices
were divided by the stock prices 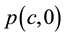 at January 4, 2010 to furnish the relative price index for each stock,
at January 4, 2010 to furnish the relative price index for each stock,
 (1)
(1)
for each trading day or trading decision d (see Figure 1). Bourse indices 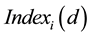 were calculated as the average of
were calculated as the average of 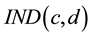 (Index in Figure 1 and Figure 2) for all 7 companies (c) in each market.
(Index in Figure 1 and Figure 2) for all 7 companies (c) in each market.
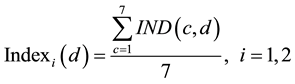 (2)
(2)
Stock volatility (vol), shown in right bottom graphic, as was calculated as the ratio (SD/MV) between standard deviation (SD) and mean (MV) of price variation between subsequent trading decisions.
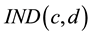 was used to calculate the current trading stock value for 50 trading decisions in M1 and M2 by multiplying the corresponding
was used to calculate the current trading stock value for 50 trading decisions in M1 and M2 by multiplying the corresponding 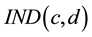 by the stock value
by the stock value 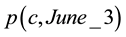 on June 3, 2012. The experiments were done on June, 3, 4 and 5. M1 is bull market with prices steadily increasing and having low volatility and M2 correspond to a high volatile transition from M1 toward a bear market that result in heavily losses in August, 2011.
on June 3, 2012. The experiments were done on June, 3, 4 and 5. M1 is bull market with prices steadily increasing and having low volatility and M2 correspond to a high volatile transition from M1 toward a bear market that result in heavily losses in August, 2011.
Relative stock prices were used to calculate stock price, by multiplying it by the value of the stocks at Portuguese Stock Market on June 2, 2012. Therefore, M1 and M2 portfolios have different values, but volunteers were restricted to 50.000 euros to trade on each market. The experiment was done from June 3 to 5, 2012.
Volunteers were divided in two groups (G1 and G2) each with 20 participants. Group 1 (10 male and 10 female) first traded in market 1 (50 decisions) and later in market 2 (another 50 decisions) while experimental group 2 (10 male and 10 female) began trading (50 decisions) in market 2 and then moved to market 1 (50 decisions).
Trading simulation progressed as follows. With the EGG mechanism in the brain, the volunteer digitized number and price of stock to trade for one and just one company and selected trading option V or C and pressed OK in order to sell or buy, respectively; or just pressed OK to maintain portfolio unaltered (see Figure 1). If price offer was within 5% variation of the next stock price, offer was accepted and the corresponding number of stock adjusted; otherwise, offer was rejected and the corresponding number of stock was maintained unaltered.
After OK was pressed a new screen was presented for another trading simulation. This new screen showed updated information of the experimental variables.
Figure 2 describes the way market 1 and market 2 behaves across the experiments and also the volatility of each stock. Is also important to say that none of the volunteer had any information about the way each of the stock will behave across the investment simulation process. Is possible to see from that Figure 2 that market 1 is a market where the stock indices decrease essential after investment decision 40 until 50 and in the market 2 the return is always growing from the first decision a until the last decision. The idea of use two different markets is to capture eventually different investment decisions behaviors in men and women when the market is growing or decreasing.
The electroencephalogram (EEG) was recorded played the game, using 10/20 protocol (impedance below 10 Kohm; low-pass filter 50 Hz, sampling frequency of 256 Hz and 10 bits of resolution). EEG epochs of 2 seconds preceding the OK button pressing. Low Resolution Electromagnetic Tomography (sLORETA) was used to spatially locate the sources  generating the recorded EEG epochs [13] [14] .
generating the recorded EEG epochs [13] [14] .
3. Results
3.1. Behavioral Data
Table 1 describes the percentage of investment decisions (buy, sell or maintain) adopted by male and female, and the percentage of rejected operations. It is important to say that the software game was formatted to avoid inadequate decisions. If price offer was within 5% variation of the next stock price, offer was accepted and the corresponding number of stock adjusted; otherwise, offer was rejected and the corresponding number of stock was maintained unaltered. The number of refused decisions made by women and men was very low, meaning that volunteers made investment decisions respecting the rules of the game.
Final Balance (Fb(d) = Portfolio value(d) + cash(d)) steadily increases from decision 1 to 100 for both male and females. The following regressions were calculated:
![]()
Figure 1. The game screen IND―the relative stock price; VAR―difference between actual and previous relative stock price, value―actual real stock price, Qt―quantity of owned stocks, total―total invested in each stock; T-Qt―proposed number of stocks to trade; price―proposed transaction price; S―selling option; B―buying option; OK―to finish proposal. Fb―Pv + C ; Pv―actual portfolio value; C―revenue (gain or loss) and A―available money for new tradings.
![]()
Figure 2. Markets M1 and M2: evolution and volatility.
![]()
Table 1. Percentage of investment decisions by gender.
![]()
R2 for these regressions were 0.74 and 0.79, respectively. Although the angular coefficient was greater for females (171) than males (117), gains were not statistically significant at level of p < 0.5.
3.2. EEG
A total of 377 (male) and 399 (female) possible sources ![]() of the Event Related Activity (2 seconds of averaged EEG prior to decision making were identified in 36 (male) and 45 (female) different cortical locations
of the Event Related Activity (2 seconds of averaged EEG prior to decision making were identified in 36 (male) and 45 (female) different cortical locations ![]() as defined by Brodmann Area number and neural structure (Table 2).
as defined by Brodmann Area number and neural structure (Table 2).
When spatial location was analyzed taking into consideration their XYZ coordinates, 26 of these locations were similar for both male and female (C in Figure 3); 63 of them were specific for females and 82 were specific for males. Spatial location for females predominated in the left hemisphere, whereas those for males predominated at the right hemisphere. Locations that were equally found in males and females predominated at anterior frontal areas and occipital areas.
![]()
Table 2. Number of Brodmann brain areas activation by gender during the 100 simulation decisions.
![]()
Figure 3. Spatial location of the identified loreta sources. numbers identify brodmann areas.
Common sources ![]() predominated over BA 9, 11, 46 and 47 at left frontal cortex and over BA 8, 9, 10 and 11 in the right frontal cortex (Figure 3 and Table 2). Common sources
predominated over BA 9, 11, 46 and 47 at left frontal cortex and over BA 8, 9, 10 and 11 in the right frontal cortex (Figure 3 and Table 2). Common sources ![]() predominated over BA 18 and 19 at the left occipital cortex and over BA 5 and 7 in the right parietal cortex (Figure 3 and Table 2).
predominated over BA 18 and 19 at the left occipital cortex and over BA 5 and 7 in the right parietal cortex (Figure 3 and Table 2).
Sources ![]() predominated over BA 8, 9, 10, 44, 46 and 47 at left frontal cortex and over BA 8, 9 and 10 in the right frontal cortex in female. In contrast,
predominated over BA 8, 9, 10, 44, 46 and 47 at left frontal cortex and over BA 8, 9 and 10 in the right frontal cortex in female. In contrast, ![]() predominated over BA 9, 10, 11, 44, 45 and 46 at left frontal cortex, and predominated over BA 8, 9, 10 and 11 in right frontal cortex in male case.
predominated over BA 9, 10, 11, 44, 45 and 46 at left frontal cortex, and predominated over BA 8, 9, 10 and 11 in right frontal cortex in male case.
Sources located around BA 4, 5 and 7 had different locations for both the left and right hemispheres and for both male and female. In the same way, sources located around BA 39 and 19 had different locations for both left and right hemispheres and for both male and female.
The most striking difference concerning gender was the predominance of sources located at the temporal cortex around BA 20, 21 and 22 mostly for the left hemisphere and females.
Being more specific about gender differences, it may be said that sources located BA1 and 2 at Postcentral Gyrus; BA 19 at Fusiform, Middle Temporal and Medial Temporal Gyri; BA at Inferior Temporal Girus; BA 32 at Anterior Cingulate Gyrus and BA at Middle Temporal and Middle Occipital Gyri were locations observed only in case of females (Table 2). In contrast, BA 6 at Precentral Gyrus; BA 19 at Inferior Occipital Gyrus and BA 46 at Middle Frontal Gyrus Anterior Cingulate Gyrus and BA at Middle Temporal and Middle Occipital Gyri were locations observed only in case of males (Table 2).
4. Discussion
Zaidi [3] and Haier et al. [4] argue that men and women use different parts of the brain to make the same kind of decisions. Present results clearly show that male and female activated different sets ![]() neurons located at different locations
neurons located at different locations ![]() to make their trading decisions.
to make their trading decisions.
Rocha et al. [15] -[18] used knowledge provided by neurosciences about neural circuits in charge of estimating benefit, risk, aversion and conflict to model decision making. This model proved to be an efficient tool to simulate stock market dynamic [5] [15] -[17] , vote decision [19] , medical diagnosing [20] , as well as moral dilemma judgment [19] .
BA 11 and medial portions of BA 9 or 10 are described in the literature as involved in benefit, risk and value assessment (e.g., [21] - [25] , while activity in middle BA 9 and 10 are reported to be associated with working memory (e.g., [21] [23] - [25] ). Here, we found sources ![]() in these cortical locations that are common to both gender, but most of the
in these cortical locations that are common to both gender, but most of the ![]() identified in these cortical areas were distinct for male and female. So, it may be proposed here that man and woman rely on different neural enrollment to estimate stock value, benefit and risk.
identified in these cortical areas were distinct for male and female. So, it may be proposed here that man and woman rely on different neural enrollment to estimate stock value, benefit and risk.
Neurons at BA 5 and 7 were proposed to be part of the neural circuits involved in arithmetic calculations [26] [27] while neurons at BA 18, 19 and 39, specially at Angular, Lingual and Fusiform Gyri were considered to be part of the neural circuits involved in arithmetic [24] [26] -[28] . Souces ![]() located 4, 5 and 7 at BA were mostly identified in the right hemisphere for both genders, whereas those located at BA 18, 19 and 39 were identified mostly in the left hemisphere. However, location sources identified at these areas differed for male and female, indicating that calculus supporting financial decision making may be gender dependent.
located 4, 5 and 7 at BA were mostly identified in the right hemisphere for both genders, whereas those located at BA 18, 19 and 39 were identified mostly in the left hemisphere. However, location sources identified at these areas differed for male and female, indicating that calculus supporting financial decision making may be gender dependent.
Finally, neurons areas located at BAs 20, 21 and 22 that are reported to contribute for recognition and semantics of numerals (e.g., [26] -[30] ). Sources located at these cortical areas were mostly identified in woman. Besides this, as pointed above, the number of ![]() s identified in the left hemisphere predominated in females a over males. One possible explanation for such a find would be to assume than female relies more on language and male relies more on numbers to reason about market conditions in order to make their decisions. For instances:
s identified in the left hemisphere predominated in females a over males. One possible explanation for such a find would be to assume than female relies more on language and male relies more on numbers to reason about market conditions in order to make their decisions. For instances:
a) Males would rely on rules such as if price of stock s is increasing at rate of ?X then buy (or sell) it, while
b) Females would rely on rules such as if price of stock s is rapidly increasing then buy (or sell) it.
5. Conclusions
Literature in finance and neurosciences describes that, on average, women are more risk averse than men when making financial decision investment. Becker et al. [2] defend that males are more likely than females to engage in risky behaviors and that gender differences are due, at least in part, to sex differences in the organization of the neural systems responsible for motivation.
Here, it is shown that gender differences on financial decision are due to distinct ways male and female recruit neurons in their brain to analyze financial date in order to make a decision. However, despite using different neural circuits to trade in a simulated stock market, both male and female were equally successful in earning above the market evolution. As far as we know, this is the first paper to provide this type of information that may be very useful in any theoretical modeling of financial market. But we have to add a word of caution, because EEG allows us to study just cortical activity, and our conclusions have to be taken into consideration under this kind of restriction.
Ethical Aspects
The authors declare that they have no conflict of interests.
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964
Helsinki
declaration and its later amendments or comparable ethical standards.
Informed consent was obtained from all individual participants included in the study.
Acknowledgements
This work was partially supported by grants LIM01-HCFMUSP, CNPq and FAPESP.