Zero-Point Field, QED Coherence, Living Systems and Biophotons Emission ()
1. Introduction
Living organisms are characterized by a high level of order and organization in order to carry out all the primitive and superior functions and actions defining life. The “structural” approach of molecular biology correlates biological properties of living matter to specific molecular species it contains, not found within non-living world as, for example, organic (macro)molecules (like DNA and RNA), proteins, polysaccharides, and other smaller molecules or inorganic ions even present in inorganic matter. This picture is analogous to those developed within the paradigm of the atomic and molecular physics in which the physical and chemical properties of materials are related to the structure and properties of atoms and molecules they contain. This is one of the most important differences between living and non-living matter. The former, in order to properly functioning, needs the prompt and continuous contribution of energy capable to generate mechanical work when necessary. From an energetic point of view, this means these organisms are able to produce and accumulate low entropy energy or what is often so-called “negentropy”, defined by Schrodinger [1] .
This is in principle possible through the Principle of Stable Non-Equilibrium (PSNE), firstly formulated by Bauer [2] , according to which: “All and only living systems are never in equilibrium. At the expense of their free energy, they ceaselessly perform work against sliding toward equilibrium demanded by the physical and chemical laws appropriate to the actual external conditions”. Living systems can then accumulate and use their energetic internal reservoirs because they are able to maintain themselves in a non-equilibrium state repre-sent- ing a sort of excited state above the ground state and whose energetic transitions produce the work needed to perform all the “living” activities. In any case no living function can be performed without an energy influx or supply to the system able to be transformed into some useful form.
According to the currently accepted interpretation of bioenergetics [3] , the fundamental external energy support is constituted by sunlight that is ultimately accumulated in organic compounds in the form of loosely bound electrons. This energy is then successively made available to the living organism as the result of an oxidation process, ending in the ATP synthesis in mitochondria and its turning into useful work.
Nevertheless, the fundamental dynamics related to ATP energy storing as free energy and its conversion into mechanical work is still somehow mysterious. In particular it must exist a mechanism able to maintain, as long as possible, the non equilibrium excited-state restoring the energy transferred to ground state converted into internal work (namely the work internally used by the system to preserve itself from going towards the equilibrium state). This work is substantially different from that performed by inanimate systems since the latter act as simple “converters” of the energy they receive from external sources to mechanical work.
Furthermore the energy restoration process requires a certain quantity of external work to be performed by a living organism, since a living system, despite the high grade efficiency of the internal work performing, gradually consumes its free energy and mass reservoir that need to be replaced with new matter from the environment. This requires an influx of energy from the “environment” (outside) towards the organism (inside) that defines the living system as “open” system from a thermodynamic point of view.
Within the context of Bauer’s theory [2] , this so called “Basic Process” then corresponds to a dynamical equilibrium in which some parts of the system leave the non-equilibrium state (so energetically “dying”) while other ones are excited acquiring an higher energy level (so making available more loosely bound electrons), increasing the free energy content as a whole and the non-equilibrium feature of the considered living state.
From a thermodynamic standpoint, it corresponds to the transition of the system from a state A, in which the internal energy is distributed among a certain number of degrees of freedom, to a new state B in which some part of this energy is transferred to and concentrated in a fewer number of such degrees of freedom. Although a process like this would seem to violate the second law of thermodynamics, in principle this is not the case as it can be proved within the framework of non-equilibrium thermodynamics of dissipative structure of self organized systems formulated by Prigogine [4] .
This energy concentration process is in fact allowed by the spontaneous statistical fluctuations of the system by which the energy density can increase in some parts at the expense of its value in other parts of the organism as often even occurs in similar processes in the inanimate world.
Nevertheless, when occurring in living systems, these fluctuations show a very important difference with respect to the inanimate world, since in the former case they are not limited in time but stay for all the length of life. Despite the several attempts to build a dynamical model able to explain how living organisms succeed to generate and store this type of highly “structured” energy, none of the commonly accepted proposals gives an ultimate physical understanding of the process, often limiting themselves to a partial theoretical modeling or to a purely phenomenological description.
In fact, in both Prigogine’s model of “dissipating structures” [4] and Bauer’s model of “Basic Process” [2] the physical origin and the governing dynamics of the parameters allowing negentropy of life still remain unknown as well as the mechanism, able to transform free energy into useful work, needed to restore the low-entropy internal energy stock.
In this paper it is proposed, basing upon a model of Quantum Vacuum (QV) dynamics already developed in previous works [5] -[7] integrated and reinterpreted within the theoretical framework of QED coherence in condensed matter [8] , a suitable mechanism able to explain the physical origin of internal energy reservoir of living organisms as the result of the dynamics between living matter and QED Zero-Point Field. Furthermore, it will show that the proposed model is also able to interpret the results of some important experimental evidences regarding the emission of biophotons by living systems under the influence of external stimuli.
2. Bioenergetics Considerations
Within the theory of “dissipative structures” [4] , it has been shown that, for a system far from equilibrium as we have seen must be a living organism, beyond the instability of thermodynamics branch that would drive the system towards an equilibrium state (i.e. the death for a living organism) and under the appropriate conditions, a new time-space behavior can emerge, characterized by the so-called “dissipative structures” [4] that can be maintained in a “stable” state only if there is a sufficient inflow of energy and matter from “outside” the system. These structures are dynamically different from the “equilibrium structures” based upon Boltzmann’s order principle. In fact, according to the latter, the entropy 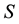 is considered as a measure of the “disorganization” of a system through the known equation
is considered as a measure of the “disorganization” of a system through the known equation
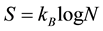 (1)
(1)
where 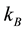 is the Boltzmann’s constant and
is the Boltzmann’s constant and 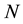 is the number of microstates associated to a given macrostate. So, stating the Equation (1), the second law of thermodynamics of the increase of entropy in an isolated system dictates the maximum level of “de-structuration” that characterizes the equilibrium state.
is the number of microstates associated to a given macrostate. So, stating the Equation (1), the second law of thermodynamics of the increase of entropy in an isolated system dictates the maximum level of “de-structuration” that characterizes the equilibrium state.
In general, if we consider a closed (but not isolated) system at a given absolute temperature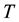 , we can define the so-called free energy function of the system
, we can define the so-called free energy function of the system 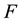 (representing, in few words, the energy convertible into useful work) as
(representing, in few words, the energy convertible into useful work) as
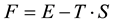 (2)
(2)
where 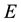 is the energy and
is the energy and 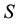 the entropy. In a non-dissipative system, the free energy is at minimum in correspondence to the equilibrium since
the entropy. In a non-dissipative system, the free energy is at minimum in correspondence to the equilibrium since 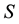 reaches its maximum value as a consequence of Boltzmann’s order principle.
reaches its maximum value as a consequence of Boltzmann’s order principle.
According to the first law of thermodynamics, for a generic transformation from a state 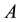 to a state
to a state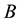 , at constant temperature (a condition almost always verified in the living systems), we can write, since both
, at constant temperature (a condition almost always verified in the living systems), we can write, since both 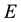 and
and 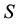 are state functions
are state functions
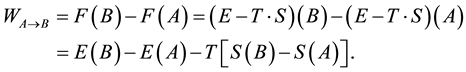 (3)
(3)
where 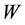 is the work exchanged by the system with the “environment” during the transformation. When we refer to living systems, as we have seen, we are interested in the processes able to produce work towards the environment, namely those for which
is the work exchanged by the system with the “environment” during the transformation. When we refer to living systems, as we have seen, we are interested in the processes able to produce work towards the environment, namely those for which![]() .
.
By Equation (3) we can see that the condition ![]() is satisfied:
is satisfied:
a) if![]() , when
, when![]() . This corresponds to an energy increase of the system that spends a fraction of its own energy just equal to
. This corresponds to an energy increase of the system that spends a fraction of its own energy just equal to ![]() to perform the transformation and produce work on the environment. This is the case of inanimate world in which the system must absorb energy from the outside and transforms a part of this into mechanical work, while the remaining part is dissipated and heats the system;
to perform the transformation and produce work on the environment. This is the case of inanimate world in which the system must absorb energy from the outside and transforms a part of this into mechanical work, while the remaining part is dissipated and heats the system;
b) if![]() , when
, when ![]() as prescribed by the second law of thermodynamics. In this case the work is done by the system towards outside environment at the expense of the positive quantity
as prescribed by the second law of thermodynamics. In this case the work is done by the system towards outside environment at the expense of the positive quantity ![]() and by decreasing its internal energy stock. This is just the case of living systems in which, as previously discussed, the non equilibrium thermodynamics permits the occurrence of fluctuations able to locally reduce the entropy, producing useful work without absorbing energy from the outside environment.
and by decreasing its internal energy stock. This is just the case of living systems in which, as previously discussed, the non equilibrium thermodynamics permits the occurrence of fluctuations able to locally reduce the entropy, producing useful work without absorbing energy from the outside environment.
Within this picture, living processes can be considered as cyclic mechanisms in which internal energy is used to produce useful work partially used to restore the internal energy stock. To ensure these processes to be stationary in order to guarantee the living functions, it is necessary the organism continuously restores its own internal high-grade energy and mass stocks.
But what could be the physical origin of this energy and mass reservoir?
3. The Emergence of Mass and Energy from QV and QED Coherence in Living Matter
It is a well-known phenomenon that the energy of a Hydrogen atom varies as a consequence of the coupling of the electric current associated to the orbiting electron to the electromagnetic field of the QV fluctuations. This effect called “Lamb-shift”, discovered in 1945 and only later understood, proves, together with other experimental evidences (as the Casimir effect and the radiative correction of the particles masses), the direct interaction between QV and atoms and that this interaction is able to modify the energy of the latter even meaningfully. In modern physics, in fact, the physical vacuum cannot be considered, due to Heisenberg uncertainty principle, as a “void” but as a physical entity manifesting a complex and fundamental background activity in which, even in the absence of matter, processes like virtual particle pair creation―annihilation and electromagnetic fields fluctuations, known as Zero Point Field (ZPF) or QV fluctuations, continuously occur.
In a model of QV recently proposed by the author [5] -[7] , both mass-less and massive particles are originated as consequence of the variation of energy density of such type of electromagnetic QV. In particular, basing on previous results, inertial and gravitational mass ![]() of a body can be generally expressed as
of a body can be generally expressed as
![]() (4)
(4)
in which ![]() is the angular frequency of ZPF mode,
is the angular frequency of ZPF mode, ![]() is the spectral energy density of QV ZPF fluctuations and
is the spectral energy density of QV ZPF fluctuations and ![]() is a function that would quantify the fraction of ZPF energy density that electromagnetically interacts with the particles contained in the “useful volume”
is a function that would quantify the fraction of ZPF energy density that electromagnetically interacts with the particles contained in the “useful volume”![]() .
.
According to this picture, we can then consider that every particle is made out of electromagnetic energy of QV and so it consists of diminished energy density of an ideal QV. For mass-less particles, the variation of energy density, corresponding to the “creation” of a particle of energy![]() , is given by
, is given by
![]() (5)
(5)
where ![]() is the QV energy density after the “extraction” of the mass-less particle. For a massive particle of rest mass
is the QV energy density after the “extraction” of the mass-less particle. For a massive particle of rest mass ![]() we have instead
we have instead
![]() (6)
(6)
where ![]() is the “proper” volume of the body and the energy density variation is considered to be concentrated, for our purpose in this paper, in the center of mass of body. From Equation (6) it immediately follows that mass can be expressed as a result of the variation of energy density of an “electromagnetic” QV
is the “proper” volume of the body and the energy density variation is considered to be concentrated, for our purpose in this paper, in the center of mass of body. From Equation (6) it immediately follows that mass can be expressed as a result of the variation of energy density of an “electromagnetic” QV
![]() (7)
(7)
or equivalently
![]() (8)
(8)
having defined![]() , and the energy of which particles are made up comes from QV.
, and the energy of which particles are made up comes from QV.
The particles composing a living organism can then interact with QV exchanging with it some amounts of energy. Living organisms could then use energy of QV to maintain lower entropy from its surrounding environment. In particular, they could reduce their internal energy stock by releasing an energy amount to QV in so reducing their entropy and performing the external work required to maintain the living functions, according to the dynamics of point b) of the above discussion. The resulting external work could be then partially used to restore the energy content of QV, maintaining the non-equilibrium state.
This energy flow from and to QV should be related, according to Equation (8), respectively to an increase and a decrease of energy and mass stock of the living system that would not be on average macroscopically detectable due to the system fluctuations as also described by the “dissipative-structures” model but could be measurable when the non equilibrium regime is perturbed by a critical external stimulus or when it slip towards an equilibrium state of low-grade energy (for example after death).
The above-proposed model can be dynamically reformulated and reinterpreted within the framework of QED coherence in condensed matter, originally developed by Preparata [8] and applied to living systems by Preparata, Del Giudice et al. [9] -[17] , according to which, basing on a well-known behavior of electromagnetic and matter quantum fields, under suitable boundary conditions (almost always verified in the condensed matter and living organisms as well), a coherent electromagnetic field, oscillating in tune with all the matter constituents, spontaneously emerges from the self-produced electromagnetic field.
In particular, it has been shown [8] that, above a critical density ![]() and below a threshold temperature
and below a threshold temperature
![]() , an ensemble of atoms or molecules, placed in the empty space (namely a volume without any matter or radiation field different than ZPF inside it), spontaneous “decays” into a more stable state (characterized by lower energy and so strongly favored) in which all the matter components are phase correlated among them by means of the action of an electromagnetic field oscillating in tune with them too, confined within a defined spatial region, called “Coherence Domain” (CD), corresponding to the wavelength of the tuning electromagnetic field.
, an ensemble of atoms or molecules, placed in the empty space (namely a volume without any matter or radiation field different than ZPF inside it), spontaneous “decays” into a more stable state (characterized by lower energy and so strongly favored) in which all the matter components are phase correlated among them by means of the action of an electromagnetic field oscillating in tune with them too, confined within a defined spatial region, called “Coherence Domain” (CD), corresponding to the wavelength of the tuning electromagnetic field.
The arising of this physical coherent state can be understood by considering that, according to quantum field theory, matter and fields continuously perform quantum fluctuations. The same types of fluctuations also characterize, as seen above, the QED QV.
We consider the matter system to be composed of electrical charged particles (electrons and nuclei) characterized by an energy spectrum ![]() and indicate with “ 0” its fundamental state (whose energy is
and indicate with “ 0” its fundamental state (whose energy is![]() ) and with “
) and with “![]() ” a generic excited state (with an associated energy
” a generic excited state (with an associated energy![]() ). A vacuum fluctuation able to coupled to the systems and excite the state
). A vacuum fluctuation able to coupled to the systems and excite the state ![]() (from fundamental one) must be have a wavelength
(from fundamental one) must be have a wavelength ![]() where
where![]() . The probability of this coupling with the excitation of state
. The probability of this coupling with the excitation of state ![]() is quantified by the “oscillator’s strength” for the transition
is quantified by the “oscillator’s strength” for the transition![]() , given by [8]
, given by [8]
![]() (9)
(9)
where ![]() is the frequency of the exciting electromagnetic field,
is the frequency of the exciting electromagnetic field, ![]() the electron mass and
the electron mass and ![]() the electromagnetic current density operator. For an atom or molecule with
the electromagnetic current density operator. For an atom or molecule with ![]() electrons,
electrons, ![]() must follow the rule
must follow the rule
![]() (10)
(10)
Now let’s consider the space volume ![]() “covered” by an oscillation of the QV electromagnetic field resonating within it, supposing it contains
“covered” by an oscillation of the QV electromagnetic field resonating within it, supposing it contains ![]() atomic or molecular species, and let be
atomic or molecular species, and let be ![]() the “Lamb-shift type” probability that a photon “escapes” from QV, couples with an atom or molecule and puts it in a given excited state. The overall probability of coupling for the
the “Lamb-shift type” probability that a photon “escapes” from QV, couples with an atom or molecule and puts it in a given excited state. The overall probability of coupling for the ![]() constituents is then
constituents is then
![]() (11)
(11)
that is proportional to the matter density. So, when density exceeds a particularly high value, almost every ZPF fluctuation couple with the atoms or molecules in the ensemble.
This condition starts the “runaway” of the system from the perturbative ground state (PGS), in which matter and quantum fluctuations are uncoupled and no tuning electromagnetic field exists, to a coherent ground state (CGS) in which, within a CD, a coherent electromagnetic field oscillates in phase with matter determining a macroscopic quantum state in which atoms and molecules lose their individuality to become part of a whole electromagnetic field + matter entangled system. The system evolution can be characterized mathematically [9] - [15] considering, for simplicity, a two-levels matter system described by the matter field ![]() with
with![]() ,
, ![]() and an electromagnetic field characterized by its vector potential
and an electromagnetic field characterized by its vector potential![]() . If we neglect the spatial dependence of both the fields (since they can be assumed slowly varying within the CD) the dynamic equations, describing the time-evolution of the electromagnetic field+ matter interacting ensemble, are given by
. If we neglect the spatial dependence of both the fields (since they can be assumed slowly varying within the CD) the dynamic equations, describing the time-evolution of the electromagnetic field+ matter interacting ensemble, are given by
![]() (12)
(12)
where
![]() (13)
(13)
![]() (14)
(14)
being ![]() the directional averaged vector potential and
the directional averaged vector potential and![]() .
.
It is easy to show [8] [17] the differential system (12) admits the following constants of motion
![]() (15)
(15)
![]() (16)
(16)
![]() (17)
(17)
in which the quantity ![]() can be considered as the “momentum” of the system and
can be considered as the “momentum” of the system and ![]() its Hamiltonian divided by
its Hamiltonian divided by![]() .
.
In order to study the time-evolution of the system we start from the “perturbative” initial state of QED defined by
![]() (18)
(18)
from which the system will “decay” towards the coherent stable state characterized by ![]() and
and![]() .
.
The short-time behavior of the system can be studied [8] by differentiating the third of Equations (12) and substituting it into the second one, so obtaining
![]() (19)
(19)
whose algebraic associated equation is
![]() (20)
(20)
As know from the general theory, the Equation (19) will have exactly three solutions (real or complex). The “decay” towards the coherent state will occur when the values of ![]() and
and ![]() are such to have only one real solution of Equation (20), the other two complex-conjugate ones just describing the exponential increase of
are such to have only one real solution of Equation (20), the other two complex-conjugate ones just describing the exponential increase of ![]() able to overcome its nearly-zero initial value and create the coherent tuning field. It can be shown [8] that this occurs, for a given
able to overcome its nearly-zero initial value and create the coherent tuning field. It can be shown [8] that this occurs, for a given![]() , when
, when
![]() (21)
(21)
where
![]() (22)
(22)
In summary, when the condition given by Equation (22) is satisfied, the system will undergo a truly “phase transition” from the PGS in which the electromagnetic and matter fields perform Zero-Point very weak uncoupled fluctuations only, towards the CGS in which a strong electromagnetic field arises from QV and couples with the oscillations of the matter fields tuning all the matter constituents to oscillate in phase with it and among themselves. But why does the system “decide” to run away towards CGS?
The answer is, as above anticipated, this state is energetically favored so representing the “true” ground state of the electromagnetic field + matter system. This can be rigorously demonstrated by mathematics [8] -[17] , but a simple physical argument can be used to justify it as follows.
If we indicate as ![]() a spontaneous QED QV fluctuation able to excite some atomic/molecular level of the matter constituents of the given ensemble and as
a spontaneous QED QV fluctuation able to excite some atomic/molecular level of the matter constituents of the given ensemble and as ![]() the energy shift induced by the interaction with the electromagnetic field of Zero Point (i.e. a Lamb-shift type term), the total energy fluctuation is given by
the energy shift induced by the interaction with the electromagnetic field of Zero Point (i.e. a Lamb-shift type term), the total energy fluctuation is given by
![]() (23)
(23)
where the minus sign before ![]() is due to the fact the Lamb-shift term reduces the energy of atomic/molecu- lar constituent, since it introduces, in the atomic Hamiltonian, the interaction term
is due to the fact the Lamb-shift term reduces the energy of atomic/molecu- lar constituent, since it introduces, in the atomic Hamiltonian, the interaction term ![]() with
with
![]() (24)
(24)
where ![]() is the atomic number and
is the atomic number and ![]() is the momentum operator of the l-th electron.
is the momentum operator of the l-th electron.
It can be shown [14] that, for an ensemble of ![]() atoms/particles interacting with ZPF, we have
atoms/particles interacting with ZPF, we have ![]() while
while ![]() so we can write
so we can write
![]() (25)
(25)
in which ![]() and
and ![]() are two constants of proportionality depending on the system properties.
are two constants of proportionality depending on the system properties.
From Equation (24) we see that it exists a definite value of![]() , depending on
, depending on ![]() and
and![]() , such that, when
, such that, when ![]() (namely just the condition for the run away of the system towards the CGS), we have
(namely just the condition for the run away of the system towards the CGS), we have
![]() (26)
(26)
The result given by Equation (26) has a very deep physical meaning since it implies some remarkable consequences [8] -[17] :
a) the CGS is the “true” ground state of the system since its energy is lower, of the quantity ![]() (gap), than the energy of “gas-like” PGS in which we only have the independent Zero-Point fluctuations of electromagnetic and matter components while, in the CGS, the matter constituents oscillates in tune with a non fluctuating “strong” electromagnetic field;
(gap), than the energy of “gas-like” PGS in which we only have the independent Zero-Point fluctuations of electromagnetic and matter components while, in the CGS, the matter constituents oscillates in tune with a non fluctuating “strong” electromagnetic field;
b) the “decay” of the system from PGS to CGS can be considered as a true phase transition, corresponding to the release of a quantity of energy just equal to the gap ![]() to the environment, so characterizing the electromagnetic field + matter ensemble as an open system;
to the environment, so characterizing the electromagnetic field + matter ensemble as an open system;
c) the tuning of the electromagnetic field with the matter field determines a renormalization of frequencies of the matter system so that the common oscillation frequency of electromagnetic field and matter field is given by![]() , where
, where ![]() is the frequency of the QV fluctuating electromagnetic field able to excite the level k and whose wavelength
is the frequency of the QV fluctuating electromagnetic field able to excite the level k and whose wavelength ![]() defines the spatial extension of CD. This implies the photon “mass- term”
defines the spatial extension of CD. This implies the photon “mass- term”![]() , namely the photons of coherent electromagnetic field are “trapped” inside the CD that so acts like a cavity. This guaranties the stability of the CGS against the irradiation of the coherent electromagnetic field towards the “outside” environment;
, namely the photons of coherent electromagnetic field are “trapped” inside the CD that so acts like a cavity. This guaranties the stability of the CGS against the irradiation of the coherent electromagnetic field towards the “outside” environment;
d) the coherent electromagnetic field generated inside a CD shows an evanescent tail ![]() (where
(where ![]() is the “radius” of a CD) at its boundary, determining a superposition between the “inner” electromagnetic fields associated to neighboring CDs. This superposition makes it possible the interaction between different CDs giving rise to the coherence among them also known as “super coherence”. This explains the physical origin of long-range and stable correlation between a very high number of matter components in living organisms;
is the “radius” of a CD) at its boundary, determining a superposition between the “inner” electromagnetic fields associated to neighboring CDs. This superposition makes it possible the interaction between different CDs giving rise to the coherence among them also known as “super coherence”. This explains the physical origin of long-range and stable correlation between a very high number of matter components in living organisms;
e) in the coherent state so far analyzed, the tuned oscillation between matter and electromagnetic field forbids any thermal fluctuation and then it is virtually associated with a thermodynamic absolute temperature![]() . In this condition, no energy inflow from the environment is then possible. This is prevented by the energy gap characterizing the coherent state after the release of the energy
. In this condition, no energy inflow from the environment is then possible. This is prevented by the energy gap characterizing the coherent state after the release of the energy![]() , representing the latent heat of phase transition. Nevertheless, if the temperature of the environment increases to a value
, representing the latent heat of phase transition. Nevertheless, if the temperature of the environment increases to a value ![]() (a CD placed in a thermal bath at
(a CD placed in a thermal bath at ![]() for example), the collisions between the fluctuating environment molecules (thermally excited) and the components of a CD, could transfer to it the energy gap per atom/molecule
for example), the collisions between the fluctuating environment molecules (thermally excited) and the components of a CD, could transfer to it the energy gap per atom/molecule![]() , able to put some of them out of tune with the electromagnetic field. This determines the “expulsion” of some matter components from the CD and the formation of an incoherent fraction of matter system at the boundaries of CD.
, able to put some of them out of tune with the electromagnetic field. This determines the “expulsion” of some matter components from the CD and the formation of an incoherent fraction of matter system at the boundaries of CD.
It is remarkable to note the formation of a CD determines a high reduction of the value of entropy inside it so “concentrating” the energy into just one degree of freedom (the matter + electromagnetic field coherent state), differently from the perturbative one in which energy can be distributed among a lot of possible microstates (the uncorrelated states of electromagnetic field and matter). This explains the physical origin of the thermodynamic condition given by Equation (3), point b), necessary in order to sustain the living functions.
As shown by the above discussion, the formation of CD is strictly related to the QV energy density dynamics: the energy needed for the generation of the coherent electromagnetic field is “extracted” from ZPF (the photons transferred from random quantum fluctuations to tuning electromagnetic field) whose energy density decrease then determines the formation and sustain of the coherent state and the release of the phase transition energy to the environment.
This more stable state “survives” at the expense of the energy supplied by ZPF fluctuations so determining a reduction of QV local energy density in the spatial region occupied by the CDs associated to a given ensemble of matter components as already assumed in the derivation of Equation (8) and supported by the dynamical considerations discussed in the following sections in relation to biophotons emission.
4. Some Experimental Evidences about Biophotons
Biophotons emission, also known as “bioluminescence”, consists in a spontaneous weak or ultraweak emission of photons by biological systems in the optical-ultraviolet frequency range whose source it is though to be related to DNA and cells activity [18] and to the functional state of a living organism [19] . The experimental studies so far performed have revealed some characterizing features of this biological emission [19] :
・ the spectral distribution ranges from![]() , namely covering the optical and the ultraviolet (UV) region of electromagnetic spectrum;
, namely covering the optical and the ultraviolet (UV) region of electromagnetic spectrum;
・ the form of the spectrum is substantially flat, i.e. without emission peaks, and follows the rule ![]() constant, where
constant, where ![]() indicates the frequency density emission;
indicates the frequency density emission;
・ the probability of counting ![]() biophotons during a given time interval
biophotons during a given time interval ![]() is given by Poisson distribution
is given by Poisson distribution![]() , where
, where ![]() is the average value of photons in the time interval
is the average value of photons in the time interval![]() , and it holds even in a non-stationary regime and for a time interval down to at least
, and it holds even in a non-stationary regime and for a time interval down to at least![]() ;
;
・ the temperature dependence of the emission doesn’t follow the Arrenius factor rather an hysteresis-like intensity curve when the temperature is cyclically increased and decreased;
・ all the observed features suggest a non-linear dependence of emission upon cell density.
Another fundamental property of biophotons emission concerns its relationship with the occurrence of an external stimulus.
For simplicity we'll consider, in this paper, the common observed behavior of biophoton signals in all the living systems when exposed to light stimulus [20] . It has been found that all biophoton signals show the similar qualitative behavior represented in Figure 1.
We can distinguish four regions:
a) pre-stimulation region 1. A continuous and almost stationary spontaneous biophotons emission is observed. It represents the pre-stimulation section of the biophoton signal or, equivalently, the asymptotic tail of the signal due to a previous stimulation;
b) stimulation region 2. This part includes the biophotons signal generated during the stimulation up to ![]() after it. In this zone the signal cannot be generally distinguished from the noise represented by the stimulus itself. In fact the intensity of stimulating visible light used in the experiments doesn’t allow the detection of the weak signal associate to biophotons emission;
after it. In this zone the signal cannot be generally distinguished from the noise represented by the stimulus itself. In fact the intensity of stimulating visible light used in the experiments doesn’t allow the detection of the weak signal associate to biophotons emission;
c) post-stimulation region 3. This is the portion of the signal emitted after light stimulation, characterized by more intense emission. It shows an exponential decay in which it firstly reduces itself by 2 - 3 orders of magnitude an then by 5% of its value within a time interval ranging from few milliseconds in humans to few minutes
![]()
Figure 1. Common qualitative behaviour of biophoton emission by living systems subjected to a light stimulus.
in photosynthetic systems. The biophotons flow intensity after the decay zone tends to the pre-stimulus value and results almost constant for a long time;
d) post-stimulation region 4. It is the post-stimulation zone, far from region 3, in which the biophoton signal asymptotically approaches the spontaneous pre-stimulation emission.
It is interesting to note that the exponential curve describing the decay of biophotonic flow after stimulation could be simply parameterized as a function of two parameters: ![]() (the total signal strength) and
(the total signal strength) and ![]() (the curve shape).
(the curve shape).
As deduced by Popp et al. [19] a careful analysis of biophoton spontaneous emissions, as well as stimulated emission in response to a stimulus (in regards to the correlation to biological processes, spectral distribution, frequency counting etc.), implies the conclusion it cannot be explained by the process of spontaneous chemiluminescence, as suggested within the commonly accepted theoretical framework of molecular biology, but requires a quantum field theory approach.
In fact, according to chemiluminescence theory, biophotons emission would be related to the inevitable tendency of living organisms to shift towards the thermal equilibrium. Nevertheless, as we have discussed in the introduction, this is not really the case for living systems since they are instead characterized, according to dissipative structures thermodynamics, by a stable non-equilibrium state associated to a low value of entropy.
Furthermore, the weak and very weak intensity of biophotons emission can be explained only in terms of high degree of coherence with a high signal to noise ratio, unexplainable by the rare and random chemiluminescence [19] .
This “classical” origin is highly improbable due to many other evidences as:
a) the strong coupling to all biological and physiological processes;
b) the non-linear optical transparency of biophotons through the matter, suggesting its highly coherent origin (the only way as we have seen, to avoid thermal dissipation inside CD);
c) the sub-Poissonian counting statistics of biophoton counting, proved by the variances lower than mean values.
As we’ll see in the following section, all these very peculiar features are simply explainable within a theoretical model based on QV energy density dynamics and a QED coherent model of condensed biological matter arising from it.
5. Discussion
From the above discussion it emerges that energy quantities lower than the gap ![]() cannot be absorbed by the single atomic/molecular component of the CD but only by the CD itself as a whole, giving rise to its excited states that are still coherent and characterized by an energy spectrum
cannot be absorbed by the single atomic/molecular component of the CD but only by the CD itself as a whole, giving rise to its excited states that are still coherent and characterized by an energy spectrum![]() . We have also seen the coherent electromagnetic field has an evanescent component extending outside the CD, able to originate [9] , on its boundary, an electromagnetic dispersive force proportional to
. We have also seen the coherent electromagnetic field has an evanescent component extending outside the CD, able to originate [9] , on its boundary, an electromagnetic dispersive force proportional to![]() . This field acts on the atoms or molecules near the CD boundary according to a force given by
. This field acts on the atoms or molecules near the CD boundary according to a force given by
![]() (27)
(27)
where ![]() is the peculiar frequency of oscillation belonging to the energy spectrum of the n-th atom or molecule,
is the peculiar frequency of oscillation belonging to the energy spectrum of the n-th atom or molecule, ![]() is the energy level of CD and
is the energy level of CD and ![]() is a damping factor.
is a damping factor.
It is remarkable to note this force acts as an electromagnetic “recognition” code, based on electromagnetic resonance, according to which the atoms or molecules characterized by an oscillation frequency ![]() are the most interacting with CD, while the sign of the force, namely its attractive o repulsive behavior, is related to the sign of the differences
are the most interacting with CD, while the sign of the force, namely its attractive o repulsive behavior, is related to the sign of the differences![]() . The atoms/molecules able to electromagnetically resonate with the CD and among each other are then “selected” by this long-range dispersive interaction and attracted to the CD boundary where they can chemically react. In this way the CD act as “enzyme” for that particular reaction, favoring the reaction between atomic/molecular resonating partners.
. The atoms/molecules able to electromagnetically resonate with the CD and among each other are then “selected” by this long-range dispersive interaction and attracted to the CD boundary where they can chemically react. In this way the CD act as “enzyme” for that particular reaction, favoring the reaction between atomic/molecular resonating partners.
This activity is particularly important in the case of water molecule that, as know, represents the basis component of every living organism. The coherent dynamics and thermodynamics of liquid water has been analyzed in series of papers [8] [12] [14] -[16] showing very peculiar and unthinkable features, whose detailed discussion can be found in the cited references. Here we recall some of these that will be used in the following analysis. It has been found that the matter system excitation involved in the water coherent phase is the transition![]() , corresponding to a
, corresponding to a ![]() electronic state of water molecule, just below the ionization threshold of
electronic state of water molecule, just below the ionization threshold of ![]() with an energy gap
with an energy gap![]() .
.
This special circumstance implies a lot part of electrons belonging to the water coherent state (actually a quantum superposition![]() ) to have a quite large probability [8] [15] to be considered as “quasi-free” electrons because of their low binding energy, so configuring the CD as an electrons donor, whereas in order to ionize non-coherent molecules high ionization energies should be required. In summary, a water CD acts, with respect its surrounding molecules, as an electromagnetic attractor, via dispersive forces, and, at the same time, as an electrons donor. Both this features confirm the role of electromagnetic “enzyme” that water molecules can play in the condensed biologic matter.
) to have a quite large probability [8] [15] to be considered as “quasi-free” electrons because of their low binding energy, so configuring the CD as an electrons donor, whereas in order to ionize non-coherent molecules high ionization energies should be required. In summary, a water CD acts, with respect its surrounding molecules, as an electromagnetic attractor, via dispersive forces, and, at the same time, as an electrons donor. Both this features confirm the role of electromagnetic “enzyme” that water molecules can play in the condensed biologic matter.
On the other hand, the role of coherence in the dynamics of enzymes, with particular reference to the availability of high-grade (low entropy) internal energy used by a living organism to respond to an external stimulus, has been already suggested by Frolich some years ago [21] .
As we have seen in the previous introduction, the reaction of a living organism to an external stimulus (or to a change of its biological state) implies the production of some mechanical work from its high-grade internal energy stock. This process can be easily explained within the QED coherent theoretical framework above outlined that can be applied not only to water but also to any other molecular species (characterized by a particular energy spectrum), provided that the conditions needed to the occurrence of coherent condensation from PGS to CGS hold (![]() and
and![]() ).
).
The process starts with the selection and attraction of the atoms/molecules, involved in the reaction chain of the response to stimulus, on the CD surface through the electromagnetic dispersive force. Then the release of a certain amount of quasi-free electrons from CD (attracted by the chemical species now present on the boundary surface of it) provides a properly enhancement of the reaction rate needed to respond to stimulus.
The loss of electrons pulls out of coherence a fraction of CD components, determining the release of photons to QV and a decrease of internal high-grade energy stock. In order to restore it, electrons can be supplied into CD from the outside, so re-establishing coherence and decreasing QV energy density. This function can be absolved by the metal co-enzymes, present in biological matter, that are able to easily transfer electrons to CD by tunnel effect. We have then the emergence of another oscillatory process in which the transition between the coherent and non-coherent state of a CD (or, at least, a part of if) continuously occurs and whose frequency is proportional to the entity of the needed response to external stimuli. It is important to note that the above process can occur not only in the fundamental energy state of a CD but also in some coherent excited states belonging to its spectrum![]() .
.
We are now in position to give an explanation of the behavior shown in Figure 1 within a quantum re-inter- pretation of the Bauer’s theory according to which living organisms use their internal energy stock to perform the external work required to respond to the external stimuli whose restoration and maintenance occur at the expense of QV energy (according the coherent―non coherent fluctuations of the “enzyme”).
This can be formally described within the proposed model of QV [5] -[7] by noting that, locally, the energy density conservation holds
![]() (28)
(28)
where ![]() is the QV energy density,
is the QV energy density, ![]() the energy density of electromagnetic radiation,
the energy density of electromagnetic radiation, ![]() is the relativistic energy density and
is the relativistic energy density and ![]() is a constant value. This can be written for, a local variation,
is a constant value. This can be written for, a local variation,
![]() (29)
(29)
or, equivalently
![]() (30)
(30)
since![]() .
.
For the transition from the PGS to CGS, Equation (30) can be written, when referred to the volume ![]() occupied by the system, as
occupied by the system, as
![]() (31)
(31)
where ![]() is the energy required to excite the matter system,
is the energy required to excite the matter system, ![]() is the energy of the coherent electromagnetic field oscillating in phase with the matter-system and
is the energy of the coherent electromagnetic field oscillating in phase with the matter-system and ![]() is the contribution to energy of the eventual interactions, but since
is the contribution to energy of the eventual interactions, but since ![]() we have, from Equation (31),
we have, from Equation (31),
![]() (32)
(32)
in agreement with the conclusions of par. 3.
If we now indicate as ![]() the overall volume associated to the living organism, we can write, by multiplying Equation (30) for the term
the overall volume associated to the living organism, we can write, by multiplying Equation (30) for the term![]() ,
,
![]() (33)
(33)
where ![]() indicates the overall mass of the living organism. Now we can put
indicates the overall mass of the living organism. Now we can put
![]() (34)
(34)
where ![]() and
and ![]() respectively are the final and initial values of mass measured at two time instants
respectively are the final and initial values of mass measured at two time instants ![]() and
and ![]() (with
(with![]() ).
).
Inserting Equation (34) into Equation (33) gives
![]() (35)
(35)
now if we put![]() , where
, where ![]() could be defined as the “QV energy mass-equivalent”, we can rewrite Equation (35) in the form
could be defined as the “QV energy mass-equivalent”, we can rewrite Equation (35) in the form
![]() (36)
(36)
Equation (36) is very interesting since it shows that living mass can be interpreted as the sum of an “initial” mass, the mass exchanged with QV (described by the term![]() ) plus a certain quantity of energy in the form of electromagnetic radiation.
) plus a certain quantity of energy in the form of electromagnetic radiation.
In order to better understand the meaning of the “living” mass terms in the previous equations we recall that, within the Bauer’s theory, the stock of structural energy that convertible to free energy ![]() able to be used for performing internal and external work, can be considered as the product of the “living substance potential”
able to be used for performing internal and external work, can be considered as the product of the “living substance potential” ![]() and the mass of “living matter”
and the mass of “living matter” ![]() (the matter residing in an excited energetic state), namely
(the matter residing in an excited energetic state), namely
![]() (37)
(37)
This equation establishes a direct relationship between free energy and living mass of an animate organism.
The experimental results above reported can now be simply interpreted within the proposed theoretical framework by assuming that living mass and energy are dynamically related to ZPF energy. This can be view reconsidering the function
![]() (38)
(38)
where![]() , obtained by Equation (35).
, obtained by Equation (35).
In the region 1, the measured value![]() , between two generic successive instant of times within the considered time interval, implies
, between two generic successive instant of times within the considered time interval, implies
![]() (39)
(39)
meaning that the variation (increase) of mass stock (and consequently of free energy by Equation (37)) of living system, necessary to maintain the system far from equilibrium, is totally compensated by a corresponding variation (decreasing) of vacuum energy density that leads to the observed stationary flow of biophotons.
In the region 2, during the light stimulus, as said above we cannot perform any type of analysis due to the masking of biophotons signal by stimulating light, but we see, from Figure 1, that at the end of this region, the value of biophotons intensity rises towards an asymptotic value (until to 5 ms from the stimulation), showing the occurrence of a phase transition, suggesting a dynamical change of QV state.
In this case we have, within this region, ![]() and, correspondingly,
and, correspondingly,
![]() (40)
(40)
meaning that the decreasing of “QV energy mass-equivalent” is greater than the “increase” of mass of the body (i.e. of the free energy stock given by Equation (37)) needed to maintain the system far from equilibrium during the stimulation, so pushing the system towards a more excited state of living matter in order to respond to the external stimulus.
This is equivalent to say that the energy supplied by QV (that decreases) must be able to furnish the free energy stock to living systems plus the radiating energy needed by the greater biophoton emission intensity. In this phase living system then “adsorbs” energy from ZPF.
This behavior has been further confirmed in a series of experiments carried out by Slawinski et al. [22] , in which a meaningful increment of biophotons production has been recorded after the different living organisms has been exposed to external stressing and lethal stimuli, so implying the condition ![]() in a volume surrounding the exposed body.
in a volume surrounding the exposed body.
In the region 3, the measured ![]() is exponentially decaying so we can write, in general,
is exponentially decaying so we can write, in general,
![]() (41)
(41)
where ![]() and
and ![]() are constant coefficients, specific of the photon-emitting unit, to be determined by fitting the experimental curves describing the biophotons emission in the different cases and the summation is done over all the possible e. m. modes of emission. In this case we obviously have
are constant coefficients, specific of the photon-emitting unit, to be determined by fitting the experimental curves describing the biophotons emission in the different cases and the summation is done over all the possible e. m. modes of emission. In this case we obviously have ![]() and then
and then
![]() (42)
(42)
The above inequality shows that now the energy density variation of QV (decrease) is not able to support the increase of living mass (i.e. of free energy) needed to preserve the non-equilibrium state so determining a decrease of biophoton intensity emission and the shift towards a less excited state of living matter, characterizing the system in the absence of the selected stimulus and suggesting a “reverse” energy flow from the living system to ZPF. Finally, in the region 4, the system behavior is dynamically equivalent to that discussed in region 1 where the tail of biophotons signal approaches the spontaneous emission rate.
In summary, according to the above model, biophotons should emitted as the result of the energy released during the transition from the non-coherent to coherent state of the “enzyme” represented by the CD of water (most probably) or of the other atomic/molecular species considered and then, ultimately, as a variation of QV energy density occurring during this transition.
This model is also able explain the main features of biophotons energy emission spectrum as, mainly, its intensity flatness and its extension from the optical to UV region.
The first aspect can be traced back to fact that, for a given molecular species and boundary conditions, the energy needed for the transition![]() , corresponding to the frequency of biophotonic quanta, is a well defined value associated to the property of a given coherent matter system (through the values of
, corresponding to the frequency of biophotonic quanta, is a well defined value associated to the property of a given coherent matter system (through the values of ![]() and
and![]() ). The second question is more complex and concerns the energetic structure of the excited state of a CD. Its details depend on the particular system we consider but, in the very representative (in regard to biological systems) case of water, it has been theoretically calculated [14] .
). The second question is more complex and concerns the energetic structure of the excited state of a CD. Its details depend on the particular system we consider but, in the very representative (in regard to biological systems) case of water, it has been theoretically calculated [14] .
The excited coherent states of water appear as coherent cold “vortex” of quasi-free electrons [14] . As recalled above, when these coherent quasi-free electrons receive an amount of energy ![]() they give rise to high number of collective, still coherent, states (with an energy spacing of the order of kHz) whose energy is given by
they give rise to high number of collective, still coherent, states (with an energy spacing of the order of kHz) whose energy is given by
![]() (43)
(43)
where ![]() is the quantized angular momentum of CD,
is the quantized angular momentum of CD, ![]() the moment of inertia of the ensemble of quasi-free electrons and
the moment of inertia of the ensemble of quasi-free electrons and ![]() an external magnetic field eventually present.
an external magnetic field eventually present.
Due to its small energy gap, water CD can only accept small amounts of energy (without escaping from coherent state) but, because of coherence, these excited states cannot thermally decay and so the remain very long living.
This means that subsequent excitations can add up, accumulating higher and higher quantities of energy whose unique limitation is represented by the value of the energy gap per molecule equal to![]() . But if we consider that a water CD averagely contains about six million molecules, the total energy stored in it easily reaches that associated to optical and UV radiation. This energy is successively released, as seen above, through a non thermal mechanism directly to the biomolecules surrounding the CD, in turn undergoing the electronic transitions in the optical and UV frequency range associated to biophotons.
. But if we consider that a water CD averagely contains about six million molecules, the total energy stored in it easily reaches that associated to optical and UV radiation. This energy is successively released, as seen above, through a non thermal mechanism directly to the biomolecules surrounding the CD, in turn undergoing the electronic transitions in the optical and UV frequency range associated to biophotons.
6. Conclusions
Living organisms continuously use their low entropy internal free energy reservoir to produce internal and external work needed to perform all the vital processes. A real understanding of how they are able to maintain this internal free energy stock, its ultimate origin and how they dynamically transform part of this energy into useful work is, despite the several models so far proposed, still substantially mysterious.
In this paper it has been suggested, basing on the theoretical framework of QED coherence in condensed matter and on a model of Quantum Vacuum recently proposed by the author in which mass and energy originate from the variable energy density of Quantum Vacuum, that living organisms could dynamically and continuously exchange energy with Quantum Vacuum, in particular “extracting” from it the energy needed to restore their own internal free energy reservoir. It has also been shown this picture is able to explain the physical origin of biophotons emission and the main features of their energy spectrum, as well as the results of a series of experiments, reporting the increase in the production of biophotons by living organisms after the exposition to more or less stressing external stimuli.
Although further studies are obviously needed in order to elaborate a complete dynamical model able to fully explain the dynamical relationship between Quantum Vacuum and living functions, the results so far obtained, demonstrate such an intimate relationship must exist.
Future researches could then open the door to the extraordinary possibility of controlling, improving or repairing biological functions by altering ZPF dynamics.
The obtained results also suggest that, in principle, the occurrence of living functions could be not strictly related, at least from an energetic standpoint, to the occurrence of the specific boundary conditions experienced on the Earth some billions of year ago, but could be associated to a more universal dynamics, driven by Quantum Vacuum. Finally, the proposed model of the Quantum Vacuum―living body interaction, if adequately improved, could offer very interesting and exciting new insights in the understanding of other still unknown processes as, first of all, human mind functioning and consciousness.