The effect of dimiristoylphosphatidylethanol on the rotational mobility of n-(9-Anthroyloxy) stearic acid in neuronal and model membranes ()
1. INTRODUCTION
The molecular mechanism of action of ethanol in the central nervous system (CNS) has long been a subject of great interest. There are several proposals for the molecular mechanism of action of ethanol, and include lipid [1-6], protein [7-13] and hydration [14-16] theories. However, the precise location of molecular action has continued to be a subject of controversy to the present day.
We have recently shown that a pathway of ethanol metabolism with an unusual phospholipid, dimyristoylphosphatidylethanol (DMPEt) increased the bulk lateral and rotational mobilities, and annular lipid fluidity of neuronal membrane lipid bilayers, and had a greater fluidizing effect on the outer monolayer [17]. We also have shown that DMPEt decreased the thickness of the neuronal membrane lipid bilayer in a dose-dependent manner with a significant decrease in thickness observed at 5 μM. There is a good possibility that the effects of ethanol on the central nervous system result not only from a direct action on neuronal membranes but also from the action of DMPEt [17]. However the fluidity of the membrane outer monolayers may vary at different positions [18,19].
Previous studies have shown that the fluorophores of anthroyloxy derivative locate at a graded series of levels from the surface to the center of the lipid bilayer structure (or a series of anthroyloxy fatty acids indicates that the depth of the group is almost linearly related to the number of carbon atoms between it and carboxyl group) [18-23]. The fluorophores of anthroyloxy derivatives can also be used to differentiate whether the bilayer has a fluidity gradient across it, as the anthrolyoxy group can be positioned at different positions of the stearic acid moiety [18-20,24-27]. These probes have been suggested to measure primarily the dynamic component of membrane fluidity [18,19,25,28-31].
The aim of this research is to provide a basis for studying the molecular mechanism of pharmacological action of DMPEt and ethanol. This study was done through investigation of the effect of DMPEt on rotational mobility of the hydrocarbon interior and polar region (membranes interface, surface region) in the native and model membrane which differ in fluidity, and was done through investigation of magnitude of differential sensitivity between native and model membrane to the fluidizing or ordering effect of DMPEt. The study was carried out using 16-(9-anthroyloxy)palmitic acid (16-AP), 12-(9-anthroyloxy)stearic acid (12-AS), 9-(9- anthroyloxy) stearic acid (9-AS), 6-(9-anthroyloxy) stearic acid (6-AS) and 2-(9-anthroyloxy)stearic acid (2-AS) those reflecting mobility at the 16, 12, 9, 6 and 2 position of aliphatic chains present in phospholipids of neuronal and model membranes.
2. MATERIALS AND METHODS
2.1. Materials
The fluorescent anthroyloxy palmitate or stearate probes, 16-AP, 12-AS, 9-AS, 6-AS and 2-AS were purchased from Molecular Probes, Inc. (Junction City, OR, USA). 1, 2-DMPC, phospholipase D, bovine serum albumin (BSA) and other reagents were obtained from Sigma (St. Louis, MO, USA) and were of analytical grade. Phospholipase D was partially purified from Savoy cabbage, using acetone precipitation, according to the method of Davidson and Long [32]. After acetone precipitation the enzyme was suspended in water and lyophilized. The powder obtained was stored at –20°C.
2.2. Synthesis of DMPEt
DMPEt was synthesized by the procedure of OmodeoSalê et al. [33], which involved the transphosphatidylation of 1, 2-DMPC/ethanol by phospholipase D. The yield was 70-80%, with the DMPEt produced purified by ion-exchange chromatography and characterized by fast atom bombardment-mass spectrometry (FAB-MS).
2.3. SPMV Preparation
The SPMV were prepared according to the procedure reported from [34,35]. The specific activities of Na, K-ATPase, acetylcholinesterase and 5'-nucleotidase in the plasma membrane fraction were approximately 4-, 2.5- and 3-times higher than those in crude homogenates. The electron microscopic examination of the prepared SPMV showed very high purity. The vesicles, which were separated according to size, demonstrated homogeneous distribution and no longer showed the presence of intracellular organelles or leakage. The protein concentration was determined by the method of Lowry et al. [36] using BSA as a standard.
2.4. Liposome Preparation
Total lipids were extracted from the SPMV as previously described [34]. Cholesterol content of the extracted total lipids was determined according to the Liebermann-Buchard reaction [37]. Phospholipids were quantitated by measuring the amounts of inorganic phosphate [38] after hydrolysis of the phospholipids at 180°C in 70% HClO4 [39]. The SPMV had a high lipid to protein ratio (0.942 mg total lipids/1 mg protein) and a low cholesterol to phospholipid molar ratio. The value was 0.593 ± 0.011 (cholesterol 0.208 ± 0.010, phospholipids 0.702 ± 0.025). An average molecular weight of 775 for phospholipids is assumed and the molecular weight of cholesterol is 387 for the calculation. Phospholipids were composed (mol%) of phosphatidylcholine (41.55 ± 0.91), phosphatidylethanolamine (36.83 ± 0.48), phosphatidylserine (13.60 ± 0.26), sphingomyelin (4.15 ± 0.16), phosphatidylinositol (2.90 ± 0.09) and lysophosphatidylcholine (0.97 ± 0.03).
The characteristics of the lipid samples, such as size, lamellarity, radius of curvature, and shape, are strongly dependent on the method used to form the vesicles [18,40,41]. As a consequence of the preparation method, the parameters that characterize the lipid phase equilibrium in lipid mixtures are affected by the lipid sample characteristics. Because the size of GUVs is on the same order as the size of cells, GUVs are becoming objects of intense scrutiny in diverse areas that focus on membrane behavior [18,41,42]. Stock solutions of total lipids or phospholipids were made in chloroform. The concentration of the lipid stock solutions was 0.2 mg/ml. Giant unilamellar vesicles (GUVs: SPMVTL or SPMVPL) with a mean diameter of 45 mm were prepared by the method developed [43-45]. To grow the GUVs, a special temperature-controlled chamber, which was previously described [41,46], was used. The experiments were carried out in the same chamber after the vesicle formation, using an inverted microscope (Axiovert35: Zeiss, Thornwood, NY). The following steps were used to prepare the GUVs: 1) ~3 µl of the lipid stock solution was spread on each Pt wire under a stream on N2. To remove the residues of organic solvent we put the chamber in a liophilizer for ~2 h. 2) To add the aqueous solvent inside the chamber (Millipore water 17.5 MW/cm), the bottom part of the chamber was sealed with a coverslip. The Millipore water was previously heated to the desired temperature (80°C for SPMVTL, 60°C for SPMVPL), and then sufficient water was added to cover the Pt wires. Just after this step, the Pt wires were connected to a function generator (Hewlett-Packard, Santa Clara, CA), and a low-frequency AC field (sinusoidal wave function with a frequency of 10 Hz and an amplitude of 3 V) was applied for 90 min. After the vesicle formation, the AC field was turned off.
2.5. Fluorescence Measurements
The fluorescence measurements were taken using a modified method of earlier study [18,19,47]. The SPMV were suspended in PBS to concentration 50 µg of protein/ml. The liposomes (SPMVTL and SPMVPL) were suspended in PBS to give a concentration 0.2 mg of total lipids or total phospholipids/ml. Stock solutions of the 16-AP, 12-AS, 9-AS, 6-AS and 2-AS in methanol (2 × 10-5 M) were prepared and kept in a cold and dark place. Aliquots were added to the solutions of the native and model membrane so that the final concentrations of the 16-AP, 12-AS, 9-AS, 6-AS and 2-AS became 4 × 10-8 M (in the case of SPMV) or 2 × 10-8 M (in the cases of SPMVTL and SPMVPL) incorporated the probes. The mixture was stirred for 20 min at room temperature in order to reduce the concentration of DMPEt that might alter the rotational mobility of the SPMV, SPMVTL and SPMVPL. Also, the mixture was bubbled by dry nitrogen for 1 min with 20 min intervals in order to eliminate oxygen that might act as a quencher. To ensure complete removal of methanol residue in the mixture, the prepared mixtures were subjected to exhausted stirring for more than 2 hr which have shown the same results as the mixtures stirred for 20 min. Concentrated solution of DMPEt was prepared in PBS and added to the labeled membrane suspension to give the desired concentration of anesthetic. The pH of the buffered sample was not changed significantly by addition of DMPEt.
Fluorescence measurements were carried out with a Multi Frequency Cross-Correlation Phase and Modulation Fluorometer (ISS K2-003), equipped with a thermostated cell holder and performed at pH 7.4 (37 ± 0.1°C). The fluorescent probes, 16-AP, 12-AS, 9-AS, 6-AS and 2-AS, were excited at 360 nm (4 nm slit width) and those emissions recorded at 445 nm (8 nm slit width) through a sharp cut-off filter (Schott KV418). Corrections for light scattering (membrane suspensions without fluorescent probes) and for fluorescence in the ambient medium (quantified by pelleting the membranes after each estimation) were made routinely, and the combined corrections were less than 9% of the total fluorescence intensity observed for anthroyloxy palmitate or stearateloaded suspensions. The intensity of the components of the fluorescence which were parallel (I∥) and perpendicular (I┴) to the direction of the vertically polarized excitation light, was determined by measuring the emitted light through polarizers oriented vertically and horizontally. Polarization (P) was obtained from intensity measurements using P = (I∥ - GI┴)/(I∥ + GI┴), where G is a grating correction factor for the monochromator's transmission efficiency for vertically and horizontally polarized light. This value is given by the ratio of the fluorescence intensities of the vertical to horizontal components when the exciting light is polarized in the horizontal direction. The polarization was expressed as anisotropy [r = 2P/(3-P)] of 16-AP, 12-AS, 9-AS, 6-AS and 2-AS.
3. RESULTS
In the present study, using fluorescence probe technique, we examined the amphiphilic effects of DMPEt on the differential rotational mobility between interface and hydrocarbon interior of SPMV, SPMVTL and SPMVPL. In order to determine effects of the DMPEt on the aforementioned rotational mobility, it was first necessary to demonstrate that the drug did not interact directly with fluorescent probes and thereby quench its fluorescence. Quenching of absorbance-corrected fluorescence intensity by the drug is not observed at all of the concentration levels where DMPEt was tested. Furthermore, if direct quenching of 16-AP, 12-AS, 9-AS, 6-AS and 2-AS by the DMPEt occurred, fluorescence lifetime would decrease. However, the fluorescence lifetime of 16-AP was not changed by the drug in the SPMV. For example, the lifetime of 16-AP in the SPMV was 11.5 ± 0.2 (n = 5), 11.1 ± 0.1(n = 5), 11.6 ± 0.3 (n = 5), 11.7 ± 0.1 (n = 5) and 11.3± 0.2 (n = 5) ns at 5, 10, 15, 20, and 25 μM DMPEt, respectively. Similar results were with 12-AS, 9-AS, 6-AS and 2-AS. Direct quenching of probe fluorescence by the drug used in the present experiments was ruled out. The anisotropy (r) values of 16-AP for hydrocarbon interior of intact SPMV, SPMVTL and SPMVPL were 0.092 ± 0.002 (n = 5), 0.070 ± 0.001 (n = 5) and 0.047± 0.001 (n = 5) at 37°C (pH 7.4) respectively (Table 1). In contrast, the values of 2-AS for interface of intact SPMV, SPMVTL and SPMVPL were 0.126 ± 0.002 (n = 5), 0.114 ± 0.001 (n = 5) and 0.096 ± 0.001 (n = 5) at 37°C (pH 7.4) respectively (Table 1). This means

Table 1. Fluorescence parameters of 16-AP, 12-AS, 9-AS, 6-AS and 2-AS in SPMV.
Fluorescence measurements were performed at 37°C (pH 7.4). Values represent the mean ± SEM of 5 determinations.
that rotational mobility of hydrocarbon interior is faster than that of membrane interface. The rotational mobility’s degrees of SPMV, SPMVTL and SPMVPL differed depending on the phospholipids, cholesterol and phospholipids are co-present and the proteins, cholesterol and phospholipids are co-present in the descending order of the SPMVPL, SPMVTL and SPMV.
3.1. Ordering Effects of DMPEt on theRotational Mobility of the MembraneInterface
The effect of the DMPEt on the anisotropy (r) of the 2-AS in the interface of SPMV, SPMVTL and SPMVPL are shown in Figures 1-3. The DMPEt increased the anisotropy (r) of the 2-AS (decreased rotational mobility) in interface of SPMV, SPMVTL and SPMVPL in a concentration-dependent manner. The significant increase in the anisotropy (r) value by DMPEt was observed even at 10 μM (Figures 1-3), respectively. The anisotropy (r) values of the 2-AS in interface of SPMV, SPMVTL and SPMVPL were raise by 0.006, 0.004 and 0.006, respectively, than those in the same region when 25 μM DMPEt was added. Variations in the anisotropy (r) values were also noticed by the change in temperature as mentioned earlier. At 37°C (pH 7.4), the anisotropy (r) of the 2-AS in interface of SPMV, SPMVTL and SPMVPL are 0.126 ± 0.002(n = 5), 0.114 ± 0.001 (n = 5) and 0.096 ± 0.001 (n = 5), respectively. On the other hand, at 25°C (pH 7.4), the anisotropy (r) of the 2-AS in interface of SPMV, SPMVTL and SPMVPL are 0.165 ± 0.003 (n = 5), 0.157 ± 0.002 (n = 5) and 0.142 ± 0.001 (n = 5), respectively. Based on the results obtained at the two different temperatures, the observed effects by the addition of 25 μM DMPEt, different values 0.006, 0.004 and 0.006 were comparable to the effect of the temperature changes by approximately 1.8, 1.1 and 1.6°C, respectively.
3.2. Disordering Effects of DMPEt on theRotational Mobility of the Hydrocarbon Interior
The effect of increasing concentrations of the DMPEt on the anisotropy (r) of the 16-AP, 12-AS, 9-AS and 6-AS in the hydrocarbon interior of SPMV, SPMVTL and SPMVPL is shown in Figures 1-3. The DMPEt decreased the anisotropy (r) of the 16-AP, 12-AS, 9-AS and 6-AS (increased rotational mobility) in a concentration-dependent manner. The significant decreases in the anisotropy (r) values of the 16-AP by the DMPEt in the SPMV, SPMVTL and SPMVPL were observed even at such low concentrations as 10 μM, respectively (Figs. 1-3). The significant decreases in the anisotropy (r) values of the 12-AS by the drug in the SPMV, SPMVTL and SPMVPL were observed even at such low concentrations as 10 μM, respectively (Figures 1-3). The significant decreases in the anisotropy (r) values of the 9-AS by the DMPEt in the SPMV, SPMVTL and SPMVPL were observed even at such low concentrations as 10, 15 and 15 μM, respectively (Figures 1-3). The DPMEt decreased the anisotropy (r) of the 6-AS in

Figure 1. The effect of DMPEt on the anisotropy (r) of the 2-AS, 6-AS, 9-AS, 12-AS and 16-AP in the SPMV. Fluorescence measurements were performed at 37°C (pH 7.4). Each point represents the mean ± SEM of 5 sample determinations. An asterisk and double asterisks signify P < 0.05 and P < 0.01, respectively, compared to control according to Student's t-test.

Figure 2. The effect of DMPEt on the anisotropy (r) of the 2-AS, 6-AS, 9-AS, 12-AS and 16-AP in the SPMVTL. Fluorescence measurements were performed at 37°C (pH 7.4). Each point represents the mean ± SEM of 5 sample determinations. An asterisk and double asterisks signify P < 0.05 and P < 0.01, respectively, compared to control according to Student's t-test.
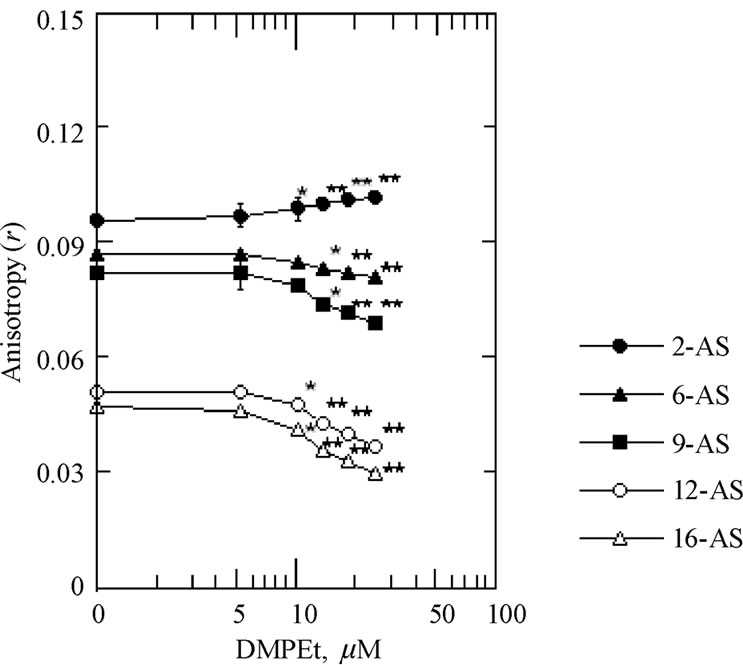
Figure 3. The effect of DMPEt on the anisotropy (r) of the 2-AS, 6-AS, 9-AS, 12-AS and 16-AP in the SPMVPL. Fluorescence measurements were performed at 37°C (pH 7.4). Each point represents the mean ± SEM of 5 sample determinations. An asterisk and double asterisks signify P < 0.05 and P < 0.01, respectively, compared to control according to Student's t-test.
a dose-dependent manner and the decrease in anisotropy (r) values in the SPMV, SPMVTL and SPMVPL by the drug was considerable at lower concentration of 10, 15 and 15 μM, respectively (Figures 1-3). The magnitude of the increased rotational mobility by the DMPEt was in the order at the position of 16, 12, 9 and 6 of aliphatic chains in phospholipids of neuronal and model membrane.
The differences in the anisotropy (r) values of the 16-AP found in hydrocarbon interior of SPMV, SPMVTL and SPMVPL before and after adding 25 μM DMPEt was 0.014, 0.014 and 0.017, respectively. These can be illustrated by comparing effects of temperature on this parameter. The anisotropy (r) of the 16-AP in hydrocarbon interior of SPMV, SPMVTL and SPMVPL are 0.092 ± 0.002 (n = 5), 0.070 ± 0.001 (n = 5) and 0.047 ± 0.001 (n = 5) at 37°C (pH 7.4), respectively. The anisotropy (r) of the 16-AP in hydrocarbon interior of SPMV, SPMVTL and SPMVPL are 0.122 ± 0.003 (n = 5), 0.106 ± 0.001 (n = 5) and 0.086 ± 0.001 (n = 5) at 25°C (pH 7.4), respectively. Thus, the differences in the anisotropy (r) values at the position of 16 in hydrocarbon interior of SPMV, SPMVTL and SPMVPL before and after adding 25 μM DMPEt was 0.014, 0.014 and 0.017, which were as large as those produced by the temperature raises of approximate 5.6, 4.7 and 5.2°C, respectively.
4. DISCUSSION
We paid a special attention to the study by Villalaín and Prieto [21] among several studies [20-24,27] in regard to 2-AS distribution in membranes. The study [21] clearly indicated the distribution region. It was reported that the 2-AS is a peculiar member of the family of probes, as the chromophore is adsorbed on the membrane interface [21]. Membrane interface is difficult to define precisely and it certainly includes the polar region near the phospholipid headgroups, or even the carbonyl groups, which are largely involved in hydrogen bonds. The degree of rotational mobility in according with the carbon numbers of phospholipids comprising of neuronal and model membranes was in the order at the position of 16, 12, 9, 6, and 2 of aliphatic chains in phospholipids, respectively. Using the membrane interface probe 2-AS, we found that DMPEt decreased the rotational mobility of lipids in the interface of neuronal and model membrane. On the contrary, using the hydrophobic interior probe 16-AP, 12-AS, 9-AS and 6-AS, we found that DMPEt increased rotational mobility of lipids in hydrophobic region of neuronal and model membrane. Furthermore, we found that in terms of increase or decrease of mobility of the neuronal membrane lipid bilayers by DMPEt, the magnitude of effects of DMPEt was greater in neuronal than model membranes.
In a dose-dependent manner, the DMPEt used in the present study lowered the anisotropy (r) values of 16-AP, 12-AS, 9-AS and 6-AS in hydrocarbon interior of SPMV but increased anisotropies of 2-AS in interface of the native and model membrane.
The DPMEt may induce disordering or ordering in their host lipids. Ordering takes place in membrane interface, whereas disordering occurs deep within the acyl chains. Although the DMPEt has strong disordering effects on hydrocarbon interior of native and model membrane, ordering effects of DMPEt on membrane interface are minor (pH 7.4, 37°C). This is consistent with the findings of Smith et al. [48] and Yun et al. [18]. This has been interpreted in terms of different locations within the bilayer for the charged and neutral forms of the anesthetic; the neutral form is located deeper within the acyl chains and consequently the DMPEt has a lesser effect on the head group [18,48].
Our data on DMPEt suggest that the observed anisotropy (r) values reflect differences in disordering or ordering constraints in hydrocarbon interior and interface of SPMV, SPMTL and SPMVPL. This is due to differences in the intrinsic component and/or the structure in interface and hydrocarbon interior of the native and model membrane. The mechanism of the action(s) of DMPEt on disordering and ordering effects on the neuronal and model membranes is not well-understood. Water associates with the head group region of phospholipids via hydrogen bonding [49]. DMPEt bind (the competitive binding of the DMPEt and water) strongly to the phosphate moiety of the phospholipids in membrane interface and weakly to the carbonyl group in competition with water, and effectively establish formation of hydrogen bonds with the carbonyl moiety, which is associated with a significant change in hydration of the DMPEt molecules themselves [50]. Incorporation of DMPEt into the native and model membranes cause alterations of the interface’s charge density of the membrane, and a conformational change in phospholipid head groups [50]. At the same time, DMPEt may exert a significant influence on hydration of the lipid bilayer. As a result, such competitive binding decreases rotational mobility and increases hydrophobicity [50]. The interaction of the DMPEt with the hydrocarbon interior will generate rearrangements of the intermolecular hydrogen-bonded network among phospholipid molecules and/or protein molecules that are associated with the liberation of hydrated water molecules on the native and model membranes [50]. The interaction will also change the orientation of the P-N dipole of phospholipid molecules [51]. These changes should cause disordering of the hydrocarbon interior, and thus they could affect the transport of Na+ and K+ in nerve membranes, leading to anesthetic action.
The sensitivities to the increasing effect of the rotational mobility of the hydrocarbon interior by the DMPEt differed depending on the native and model membranes in the descending order of the SPMV, SPMVPL and SPMVTL. When we take the results of this study and Jang’s previous study [17,52], it is without a doubt that the DMPEt increases the rotational mobility of the hydrocarbon interior of the membrane. What could be the reason that the effects on neuronal membranes where phospholipids, cholesterol and proteins are co-present are greater compared to the effect on model membranes where the protein is not co-present? It is presumed because proteins magnify the effect of DMPEt on lipids through protein-lipid interaction. These effects are not solely due to the influence of the DMPEt on lipids, but they are magnified by the interaction between lipids, proteins and water. Water plays a fundamental role in cell membrane structure in that it drives the formation of the lipid bilayer, with a polar surface facing the aqueous environment and a hydrophobic interior containing the fatty acyl chains and transmembrane proteins. In general, the structure and dynamics of proteins are also to a large extent governed by interactions with water [53]. Water penetrates into lipid bilayers at least as far as the glycerol backbone and also deeper between fatty acyl chain packing defects. Water at the proteinlipid interface is an additional factor involved in influencing the lipid bilayer structure. The introduction of small peptides, consisting of three amino acids, can cause a shift of water deeper into the bilayer, indicating increased hydration [54]. Altered hydration may have marked effects on membrane protein/lipid functioning, possibly due to the formation of hydrogen bonds between the interchain water and protein amino acid side chains facing/lipid acyl chains facing into the hydrophobic interior of the membrane. It is possible that the proteins organize the lipid in a way that makes them more susceptible to the drug.
Ethanol increased the lateral and the rotational mobilities of plasma membrane vesicles (CHOK1-PMV) of cultured Chinese hamster ovary K1 cells [5], the plasma membrane vesicles (ATCC-PMV) of cultured hybridoma cells (ATCC T1B 216) [55], plasma membrane vesicles of the cultured mouse myeloma cell line Sp2/0-Ag14 [6] and SPMV [56]. Ethanol had a greater effect on increasing the range of rotational mobility of the outer monolayer compared to the inner monolayer of CHOK1- PMV [5], ATCC-PMV [55], Sp2/0-Ag14 [6] and SPMV [56]. Furthermore, as mentioned in the introduction, a pathway for ethanol metabolism, where the product was an unusual phospholipid, PET [57], has been reported. Judging from the results of the present study, as well as those from other studies [17,33,57,58], there is a good possibility that the effects of ethanol on the CNS result not only from a direct action on neuronal membranes, but also from the action of DMPEt.